 2022, Vol. 49
2022, Vol. 49扩展功能
文章信息
- 李言, 笪心怡, 张雨晨, 方馨, 周萍萍
- LI Yan, DA Xinyi, ZHANG Yuchen, FANG Xin, ZHOU Pingping
- 酿酒酵母芳樟醇耐受性的工程改造
- Engineering of Saccharomyces cerevisiae for improved tolerance to linalool
- 微生物学通报, 2022, 49(8): 3062-3078
- Microbiology China, 2022, 49(8): 3062-3078
- DOI: 10.13344/j.microbiol.china.211198
-
文章历史
- 收稿日期: 2021-12-16
- 接受日期: 2022-04-17
- 网络首发日期: 2022-05-18

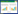




芳樟醇具有特殊的香气,是目前工业上用量最大的香精香料之一[1]。芳樟醇不仅具有抗菌、驱避杀虫、消炎止痛及镇静等多种生物学功效,还可以作为维生素A、维生素E及异植物醇生产过程中的重要中间体[2],是食品、医药和化妆品行业的重要原料[3]。目前芳樟醇主要通过植物提取和化学合成两种方式来制备[2]。由于植物提取的芳樟醇不仅产量很低,而且分离提纯步骤复杂,并不能满足人们日益增长的需求[4];而化学合成的芳樟醇是混合消旋体,无构型选择性且合成工艺复杂,成本相对高[5]。随着合成生物学技术的高速发展,越来越多的研究开始关注如何将微生物改造成细胞工厂来进行生物炼制,以达到将生物原料转化为人类所需要生化产品的目的[6-7]。近几年,随着芳樟醇合成酶的发现,代谢改造微生物生产芳樟醇成为重要选项[8-9]。
在酵母芳樟醇的生物合成研究中,Amiri等在酿酒酵母中表达了薰衣草来源的芳樟醇合成酶,并用MET3启动子下调角鲨烯合成酶(Erg9)并过表达甲羟戊酸(mevalonic acid,MVA)途径中3-羟基-3甲基-辅酶A还原酶的催化区段(tHmg1),使酿酒酵母中芳樟醇的产量达到了95 μg/L[10]。Deng等通过将软枣猕猴桃来源的芳樟醇合成酶AaLS1与法呢基焦磷酸合酶Erg20在酿酒酵母中进行融合表达,并在3 L发酵罐中进行了分批发酵,芳樟醇的产量达到240 μg/L[5]。2017年,Cao等通过在解脂耶氏酵母中过表达MVA途径中的限速酶以及构建并表达Erg20F88W-N119W突变体,使芳樟醇产量达到了6.96 mg/L[11]。最近,Peng等通过降低Erg20半衰期从而下调Erg20活性,使芳樟醇产量达到了约18 mg/L[12]。在本课题组前期的研究中,通过对芳樟醇合成酶进行定向进化筛选到了一株活性提高的突变体[13]。在此基础上,利用SKIK标签进一步提高了芳樟醇合成酶的表达量,并通过RIAD和RIDD短肽序列将t67OMcLisE343D/E352H和Erg20F96W/N127W进行了体内无支架组装,提高了芳樟醇合成酶对前体GPP的竞争效率,最后利用不同强弱启动子对酿酒酵母内源Erg20进行了下调,使芳樟醇摇瓶产量达到80.9 mg/L[14],这是目前所报道的在酿酒酵母中从头合成芳樟醇的最高产量,然而该产量远不能满足工业化生产所需。
截至目前,关于芳樟醇生物合成的研究主要集中在不同来源芳樟醇合成酶的挖掘、途径中限速酶的改造和竞争途径的下调等方面。然而,一些研究表明单萜类物质会破坏细胞膜的结构[15]、影响细胞壁的功能[16]、诱发细胞内活性氧(reactive oxygen species,ROS)的积累[17-18]及干扰胞内能量代谢[19],从而对微生物细胞造成很强的毒性[20]。因此,为了提高微生物合成芳樟醇的产量,必须提高细胞对芳樟醇的耐受性。目前关于酿酒酵母对芳樟醇耐受性机制及改造方面的研究报道甚少,但有研究者通过毒理基因组学实验揭示了烷烃类物质与酿酒酵母细胞相互作用的分子机理,在C9和C10烷烃的刺激下,与外排泵、膜修复、自由基解毒等相关的基因被上调或者下调表达[21]。在产物转运方面,ABC转运蛋白相关基因如ror1、snq2、pdr5和pdr15会上调促进产物转运到胞外,从而减少胞内有毒产品的积累[21]。在酿酒酵母中异源表达Grosmannia clavigera来源的ABC转运蛋白(GcABC-G1),有效地提高了酿酒酵母对3-蒈烯、柠檬烯和β-蒎烯的耐受性[22-23],但同时也发现,过表达ABC-G1对α-蒎烯的耐受性无影响[23]。在自由基解毒方面,抗氧化系统相关的基因skn7、yap1、yap5、stb5、gpx2和gsh2等会出现显著的差异表达,消除细胞内活性氧的积累,从而增强细胞的抗毒性能力[18, 21]。除了上述这些调控,还存在全局转录调控因子如Ino2,可以同时调控与外排泵、自由基、细胞膜修复相关的基因[21]。最近,Li等通过解脂耶氏酵母转录组学分析发现,在添加柠檬烯时82个基因上调表达,这些基因多数与膜转运、抗药性有关,其中分别过表达8个基因(YALI0F19492g、ftr、muc19、bama、wars、ndk、ant和phc)可以不同程度地降低解脂耶氏酵母在含柠檬烯培养基中的生长延滞期[22]。
除了对耐受性已知的相关基因进行靶向遗传改造外,传代适应性驯化以及常压室温等离子体诱变(atmospheric and room temperature plasma,ARTP)等技术是目前提高微生物细胞耐受性的重要方式。研究者通过酿酒酵母连续传代的自适应进化获得了柠檬烯耐受性提高的酿酒酵母突变株,并通过全基因组测序发现截短的Tcb3蛋白(tTcb3p1−989突变体,只保留Tcb3蛋白N端1−989位氨基酸)对酿酒酵母耐受性改变起了重要作用[24]。Tcb3蛋白属于tricalbin蛋白家族一员,主要定位于皮质内质网-质膜膜接触位点,具有促进皮质内质网与质膜连接并介导脂质转运的功能,缩短的tTcb3p1−989缺失了C2脂质结合构域,使得其从质膜中脱离出来并定位在核周内质网和皮质内质网中[24]。过表达tTcb3p1−989突变体对酿酒酵母细胞壁结构的维持和修复具有重要的影响,使酿酒酵母对D-柠檬烯、蒎烯和月桂烯这3种单萜物质的耐受性分别提高了9、11和8倍[24]。肖琳等通过ARTP等离子体诱变育种技术对恶臭假单胞菌进行了随机诱变,使恶臭假单胞菌对正丁醇的耐受浓度提高3倍[25]。
上述研究表明,通过对耐受性相关基因的定向操作或者对微生物细胞进行定向驯化均可提高菌株的耐受性。由于不同化合物结构有所差异,耐受性调控方面也存在很大差别[20]。本研究通过在酿酒酵母中分别过表达ABC转运蛋白(Yor1、Snq2、Pdr5、Pdr15和Pdr18)、ROS调控相关酶(Gre2、Ctt1、Yhb1、Gpx2、Trr1、Trx2和Gsh2)及转录调控因子(Ino2、Yap1、Yap5和Stb5),考察其对酿酒酵母芳樟醇耐受性的影响,并对酿酒酵母BY4741进行了传代适应性驯化,以期获得芳樟醇耐受性提高的酿酒酵母突变株,然后通过基因组重测序分析相关的基因突变位点,阐明耐受性提高的潜在分子机制。
1 材料与方法 1.1 材料大肠杆菌DH5α用于重组质粒克隆的扩增。
LB液体培养基:酵母提取物5 g/L,蛋白胨10 g/L,NaCl 10 g/L,琼脂(固体培养基加入) 15−20 g/L,用1 mol/L NaOH将pH调成7.0,0.15 MPa湿热灭菌20 min。LB培养基用于大肠杆菌的培养。
YPD (yeast extract peptone dextrose)培养基:酵母浸出粉10.0 g/L,蛋白胨20.0 g/L,葡萄糖20.0 g/L,琼脂(固体培养基加入) 15.0−20.0 g/L,在0.1 MPa条件下灭菌21 min,用于酿酒酵母的培养。
10×YNB母液:称取17.0 g/L YNB和50.0 g/L (NH4)2SO4溶于ddH2O中,用0.22 μm无菌针式过滤器过滤除菌。
SD-URA−培养基:葡萄糖20.0 g/L,10% (体积分数)的10×YNB母液,L-组氨酸20.0 mg/L,L-蛋氨酸20.0 mg/L,L-亮氨酸100.0 mg/L,琼脂(固体培养基加入) 15.0−20.0 g/L。
DNA限制性内切酶、高保真酶DNA聚合酶(Prime STARTM HS DNA Polymeras)和T4 DNA连接酶,大连宝生物(TaKaRa)公司;核酸电泳相关试剂、抗生素(氨苄青霉素、卡那霉素、G418遗传霉素)和酵母基因组抽提试剂盒,生工生物工程(上海)有股份限公司;细菌质粒抽提试剂盒、PCR产物纯化试剂盒和DNA凝胶纯化等试剂盒,爱思进生物技术(杭州)有限公司;芳樟醇标准品,上海阿拉丁生化科技股份有限公司。
实验所用引物信息见表 1,由生工生物工程(上海)股份有限公司合成。
| 引物名称 Primer name |
引物序列 Primer sequences (5′→3′) |
| PTEF1-F | CCTGCATACCCCTCATTTCCACGACCGCGAATCCTTACATCACAC |
| PTEF1-R (BamHⅠ) | CAGCGGGATCCTTTGTAATTAAAACTTAGATTAG |
| PHXT7-F | GTGTGATGTAAGGATTCGCGGTCGTGGAAATGAGGGGTATGCAGG |
| PHXT7-R (NotⅠ) | GTTGTTGCGGCCGCTTTTTGATTAAAATTAAAAA AAC |
| yor1-F (NotⅠ) | GAAGAAGCGGCCGCATGACGATTACCGTGGGG |
| yor1-R (SacⅠ) | CGGCGAGCTCTTAACTTCTGTTCTCGAAATC |
| snq2-F (NotⅠ) | GAAGAAGCGGCCGCATGAGCAATATCAAAAGCACG |
| snq2-R (SacⅠ) | CGGCGAGCTCTTACTGCTTCTTTTTCCTTATGT |
| pdr5-F (NotⅠ) | GAAGAAGCGGCCGCATGCCCGAGGCCAAGC |
| pdr5-R (SacⅠ) | CGGCGAGCTCTTATTTCTTGGAGAGTTTACCGT |
| pdr15-F (BamHⅠ) | CAGGGATCCATGTCATCAGATATCAGAGACGTAGA |
| pdr15-R (SalⅠ) | GGAAGTCGACTCACTTCTTGGGTTTTTCGG |
| pdr18-F (NotⅠ) | GAAGAAGCGGCCGCATGGAATGCGTTTCAGTAGA |
| pdr18-R (SacⅠ) | CGGCGAGCTCCTAAATGAAACCGAAGTTTCTC |
| gre2-F (BamHⅠ) | CAGGGATCCATGTCAGTTTTCGTTTCAGG |
| gre2-R (XhoⅠ) | CCTTCTCGAGTTATATTCTGCCCTCAAATT |
| ctt1-F (BamHⅠ) | CAGGGATCCATGAACGTGTTCGGTAAAAAAG |
| ctt1-R (XhoⅠ) | CCTTCTCGAGTTAATTGGCACTTGCAATGGA |
| yhb1-F (BamHⅠ) | CAGGGATCCATGCTAGCCGAAAAAACC |
| yhb1-R (XhoⅠ) | CCTTCTCGAGCTAAACTTGCACGGTTGAC |
| grx2-F (BamHⅠ) | CAGGGATCCATGACCACATCTTTTTATGAT |
| grx2-R (XhoⅠ) | CCTTCTCGAGTCATTTACTTAACAGGCTTTGG |
| trr1-F (BamHⅠ) | CAGGGATCCATGGTTCACAACAAAGTTAC |
| trr1-R (XhoⅠ) | CCTTCTCGAGCTATTCTAGGGAAGTTAAGTA |
| trx2-F (BamHⅠ) | CAGGGATCCATGGTCACTCAATTAAAATCC |
| trx2-R (XhoⅠ) | CCTTCTCGAGCTATACGTTGGAAGCAATAGC |
| gsh2-F (BamHⅠ) | CAGGGATCCATGGCACACTATCCACCT |
| gsh2-R (XhoⅠ) | CCTTCTCGAGCTAGTAAAGAATAATACTGTCC |
| ino2-F (BamHⅠ) | CAGGGATCCATGCAACAAGCAACTGGGAA |
| ino2-R (SalⅠ) | GGAAGTCGACTCAGGAATCATCCAGTATGTGC |
| yap1-F (NotⅠ) | GAAGAAGCGGCCGCATGAGTGTGTCTACCGCCAAGA |
| yap1-R (SacⅠ) | CGGCGAGCTCTTAGTTCATATGCTTATTCAAAGC |
| yap5-F (BamHⅠ) | CAGGGATCCATGGCTCTACCTCTGATAAA |
| yap5-R (XhoⅠ) | CCTTCTCGAGTCAGTGGATGATGGACCG |
| stb5-F (NotⅠ) | GAAGAAGCGGCCGCATGGATGGTCCCAATTTTG |
| stb5-R (SacⅠ) | CGGCGAGCTCTCATACAAGTTTATCAACCCA |
| tRNA-R (Esp3Ⅰ) | ATCGCGTCTCAAGCTTTGCGCAAGCCCGGAATCG |
| gYBR172C-F (ESP3Ⅰ) | ATCGCGTCTCTGCGCAATACAAATCGACCACTATGCGTTTTAGAGCTAGAAATAGCAA |
| YBR172C-HAF | TAGTCAAACAGATGAATCTCACAAGCAAGAATACAAATCGACCACTATGCTAGAAAAAGGGAAGAAGGAAAAAT |
| YBR172C-HAR | TGTGTCCTTTTTTGTCTTTTC |
| gYHR007C-F (ESP3Ⅰ) | ATCGCGTCTCTGCGCACCATACTTACCTTTCGGTGGGTTTTAGAGCTAGAAATAGCAA |
| YHR007C-HAF | AGAGACGAATACTTCCCTAA |
| YHR007C-HAR | TGGAAGAGTAACCATAGATG |
| gYMR275C-F (ESP3Ⅰ) | ATCGCGTCTCTGCGCAGCTAGATGTCACTTGTAAGCGTTTTAGAGCTAGAAATAGCAA |
| YMR275C-HAF | GTAAAAGAACGATTAAACGG |
| YMR275C-HAR | TATTTATCAATACCAAAACTGGGAGGTAACAAACAATGAGAGAAATGCTCTTGCTTACAAGTGACATCTAG |
| gYBR074W-F2 (ESP3Ⅰ) | ATCGCGTCTCTGCGCAGCATAAACTTGATTTCGATCGTTTTAGAGCTAGAAATAGCAA |
| gYBR074W-HAR3 | TCATGTTTTACATAATAACCGAAACTCAACAGAGAGATCTCCTTACGAACCAATAGTTG |
| gYBR074W-HAR3 | CCATCCGTCAAAAGTTTTGGAAACCACTGGACTCCGATATGGTAAGTTTCTTGATCGAAATCAAGTTTATG |
以PTEF1-R (BamHⅠ)和PTEF1-F为引物、酿酒酵母BY4741基因组DNA为模板,PCR扩增PTEF1启动子片段;以PHXT7-F和PHXT7-R (NotⅠ)为引物、酿酒酵母BY4741基因组DNA为模板,PCR扩增出PHXT7启动子片段;然后以PTEF1-R (BamHⅠ)和PHXT7-R (NotⅠ)为引物、PTEF1和PHXT7启动子片段为模板,通过融合PCR方法扩增出PTEF1-PHXT7双向启动子片段,将片段用BamHⅠ和NotⅠ进行双酶切后与同样双酶切的质粒pUMRI-15[26]进行连接,然后转化到大肠杆菌DH5α中,提取得到质粒pUMRI-18。
根据酿酒酵母S288C的yor1、snq2、pdr5、pdr15、pdr18、gre2、ctt1、yhb1、gpx2、trr1、trx2、gsh2、ino2、yap1、yap5和stb5的基因序列设计引物(表 1),并以酿酒酵母BY4741基因组DNA为模板PCR扩增目的基因,将上述基因片段用BamHⅠ/XhoⅠ或者NotⅠ/SacⅠ双酶切,连接到pUMRI-18质粒上,转化至大肠杆菌DH5α感受态细胞中,获得相应的pUMRI-18-yor1、pUMRI-18-snq2、pUMRI-18-pdr5、pUMRI-18-pdr15、pUMRI-18-pdr18、pUMRI-18-gre2、pUMRI-18-ctt1、pUMRI-18-yhb1、pUMRI-18-gpx2、pUMRI-18-trr1、pUMRI-18-trx2、pUMRI-18-gsh2、pUMRI-18-ino2、pUMRI-18-yap1、pUMRI-18-yap5和pUMRI-18-stb5重组质粒,将pUMRI-18及重组质粒用SfiⅠ线性化后,用醋酸锂转化方法[27]整合到酿酒酵母BY4741基因组中,获得相应的酿酒酵母工程菌株。
采用引物gYBR172C-F (Esp3Ⅰ)和tRNA-R (Esp3Ⅰ),并以M3-T质粒为模板[28] PCR扩增出gRNA序列片段,用Esp3Ⅰ进行酶切,然后与Esp3Ⅰ酶切后的TYPE3-M质粒[28]进行连接转化,构建TYPE3-M-gYBR172C质粒。然后采用YBR172C-HAF/YBR172C-HAR为引物对,以ScM6酵母基因组DNA为模板扩增出HAYBR17C同源臂序列。将构建的质粒和同源臂共转化到酿酒酵母BY4741菌株中,并涂布在SD-URA−平板上,提取酵母转化株基因组DNA,通过PCR扩增测序验证ybr172c基因序列,筛选出发生YBR172CK404Q突变的菌株,并命名为BY4741-YBR172CK404Q。同样构建BY4741-YHR007CG446R、BY4741-YMR275CF384C、BY4741-YBR074WT838N菌株及同时携含有YBR172CK404Q、YHR007CG446R、YMR275CF384C、YBR074WT838N这4个基因突变的BY4741-M4菌株。
1.3 培养条件首先将酿酒酵母菌株在YPD固体平板进行划线,放置30 ℃培养3 d后,挑取单克隆接种到5 mL YPD试管中,于30 ℃、220 r/min恒温摇床中培养12−15 h左右。然后按照0.5% (体积分数)的接种量转接到含有8 mL YPD培养基的25 mL细菌瓶中,于30 ℃、220 r/min恒温摇床中培养72 h。将芳樟醇溶于DMSO中配制成不同浓度母液,并按照需要添加40 μL不同浓度的母液至8 mL YPD培养基中。
1.4 菌株适应性传代驯化以酿酒酵母BY4741为出发菌株、以172 mg/L的芳樟醇为初始浓度进行接种、生长和传代。每隔15代左右,逐渐提高芳樟醇浓度(258、344、430、516、602和688 mg/L)进行传代适应性驯化;并将最后传代驯化后的菌液稀释,分别涂布在添加了不同芳樟醇浓度(645、860、1 075和1 290 mg/L)的YPD固体平板中。通过菌落大小初步筛选出耐受性提高的酿酒酵母突变株,并按照1.3中液体培养条件进行复筛。
1.5 全基因组重测序采用酿酒酵母基因组提取试剂盒提取酿酒酵母基因组DNA,并将提取的基因组DNA送至苏州金唯智生物科技有限公司进行二代基因组重测序分析。参考基因组为https://downloads.yeastgenome.org/sequence/strains/BY4741/。
1.6 菌株的遗传稳定性分析挑取单菌落到5 mL YPD培养基的试管中,30 ℃、220 r/min培养12 h后,取50 μL菌液转接到新的5 mL YPD试管培养基中进行传代培养,每转接一次作为一次传代,并连续进行20次传代。将第10、15和20代的培养物进行稀释涂平板,挑取单菌落,转接到5 mL YPD试管中,30 ℃培养12−15 h,然后按照0.5% (体积分数)的接种量转接至含有8 mL YPD培养基的25 mL培养瓶中,在培养基中添加430 mg/L的芳樟醇,考察传代菌株的芳樟醇耐受情况。
2 结果与分析 2.1 芳樟醇对酵母细胞生长的影响在含8 mL YPD培养基的细菌培养瓶中,分别添加107.5、215.0、322.5和430.0 mg/L的芳樟醇,并接种野生型酿酒酵母BY4741,检测其生长情况。结果如图 1所示,在培养基中添加107.5 mg/L以上的芳樟醇,酵母的生长受到了不同程度的限制;当添加322.5 mg/L的芳樟醇时,酵母的最终生物量下降了55.5%;添加430.0 mg/L的芳樟醇,酵母的生长受到了严重的抑制。上述实验证实芳樟醇对酵母细胞具有较强的毒性。

|
| 图 1 芳樟醇对酿酒酵母生长的影响 Figure 1 Growth curves of S. cerevisiae strain under different concentration of linalool. |
|
|
研究选取了5个ABC转运蛋白(Yor1、Snq2、Pdr5、Pdr15和Pdr18),将编码这些蛋白的基因置于强启动子PTEF1或者PHXT7下进行表达。为考察重组菌株对芳樟醇的耐受效果,将转化了pUMRI-18质粒的对照菌株BY4741-CK及重组菌株BY4741-yor1、BY4741-snq2、BY4741-pdr5、BY4741-pdr1515和BY4741-pdr18在含322.5 mg/L芳樟醇的YPD培养基中进行培养,并测定生长曲线。结果如图 2A所示,与对照菌株BY4741-CK相比,分别过表达Yor1、Snq2、Pdr5、Pdr15和Pdr18蛋白并不能有效提高酿酒酵母对芳樟醇的耐受性。芳樟醇作为一种小的疏水分子,可能存在另外的转运机制。有研究表明,单萜类物质会引发细胞内ROS的积累[18]。因此,本研究分别在酿酒酵母中过表达了Gre2 (甲基乙二醛还原酶)、Ctt1 (胞质过氧化氢酶)、Yhb1 (一氧化氮氧化还原酶)、Gpx2 (磷脂过氧化氢谷胱甘肽过氧化物酶)、Trr1 (胞质硫氧还蛋白还原酶)、Trx2 (硫氧还蛋白异构酶)和Gsh2 (谷胱甘肽合成酶)这7个与ROS调控有关的酶,在含322.5 mg/L芳樟醇浓度的培养基中进行培养,生长情况如图 2B所示,单独过表达这几个ROS调控相关的酶也并未提高酿酒酵母对芳樟醇的耐受性。此外,本研究还选取了Ino2 (转运、ROS调控、膜修复相关的转录调控因子)、Yap1 (转运以及ROS相关的转录调控因子)、Yap5 (ROS相关的转录调控因子)和Stb5 (ROS相关的转录调控因子)这4个转录调控因子,研究其对芳樟醇耐受性的影响。类似地,在添加了322.5 mg/L芳樟醇的培养基中进行培养并测定生长曲线。结果如图 2C所示,与对照菌株BY4741-CK相比,单独过表达这4个转录调控因子的菌株并未表现出生长优势。

|
| 图 2 过表达ABC转运蛋白、ROS调控相关酶及转录调控因子对酿酒酵母生长的影响 Figure 2 Growth curves of S. cerevisiae strains overexpressing different ABC transporters, ROS-related enzymes and transcription factors. A: The effect of individually overexpressing five ABC transporters on S. cerevisiae cells growth; B: The effect of individually overexpressing seven ROS-related enzymes on S. cerevisiae cells growth; C: The effect of individually overexpressing four transcription factors on S. cerevisiae cells growth. The medium was supplemented with 322.5 mg/L of linalool. A:分别过表达5个ABC转运蛋白对酿酒酵母生长的影响;B:分别过表达7个ROS调控相关酶对酿酒酵母生长的影响;C:分别过表达4个转录调控因子对酿酒酵母生长的影响。培养基中添加了322.5 mg/L芳樟醇 |
|
|
由于上述单个基因的过表达并未有效提高酿酒酵母芳樟醇的耐受性,因此进一步通过酿酒酵母传代驯化培养,定向获得芳樟醇耐受性提高的酿酒酵母菌株。在这个过程中,首先需要建立一套高效的筛选方法。实验发现,与添加860 mg/L芳樟醇的固体平板相比,酿酒酵母菌株在不含芳樟醇的平板中生长更快、菌落更大(图 3),因此可以利用菌落大小筛选耐受性提高的菌株。

|
| 图 3 酿酒酵母在含不同芳樟醇浓度平板中的菌落大小 Figure 3 Colony size of S. cerevisiae on plate with addition of different concentration of linalool. A: The growth of yeast on the plate without addition of linalool; B: The growth of yeast on the plate with 860 mg/L linalool addition. A:平板中不添加芳樟醇时酵母的生长情况;B:平板中添加了860 mg/L芳樟醇时酵母的生长情况 |
|
|
在驯化过程中,以酿酒酵母BY4741为出发菌株,以172 mg/L的芳樟醇为初始浓度进行试管传代培养。每隔15代左右,逐渐提高芳樟醇浓度(258、344、430、516、602和688 mg/L)进行传代适应性驯化;并将最后传代驯化后的菌液稀释,分别涂布在含有不同芳樟醇浓度(645、860、1 075和1 290 mg/L)的YPD固体平板中。通过菌落大小,初步筛选出两株长势较好的突变株ScM2和ScM6。将野生型、ScM2和ScM6菌株进行稀释点板实验。图 4结果显示,在固体培养基中添加1 075 mg/L芳樟醇时野生型已不能生长,而ScM2和ScM6在含1 290 mg/L芳樟醇的平板上还能生长。

|
| 图 4 稀释点板实验验证酿酒酵母突变株对芳樟醇的耐受能力 Figure 4 Detection tolerance of S. cerevisiae variant strains to linalool by spot dilution assays. A: Without addition of linalool; B: With addition of 860 mg/L linalool; C: With addition of 1 075 mg/L linalool; D: With addition of 1 290 mg/L linalool. 10−1−10−6 represents dilution multiple. A:不添加芳樟醇;B:添加860 mg/L芳樟醇;C:添加1 075 mg/L芳樟醇;D:添加1 290 mg/L芳樟醇。10−1−10−6表示稀释倍数 |
|
|
进一步将野生型、ScM2和ScM6菌株进行了液体培养验证。在含8 mL YPD培养基的细菌瓶中添加了0、322.5、430.0和645.0 mg/L芳樟醇,并接种这3株菌,分别培养24、48和72 h,检测OD600。结果如图 5所示,在不含芳樟醇的情况下,突变菌株ScM2和ScM6与野生株生长相似,而在添加了322.5、430.0和645.0 mg/L芳樟醇的情况下,这两株突变株生长明显较优;在含430 mg/L的芳樟醇中,野生菌株在培养48 h时几乎不生长,而突变株ScM2和ScM6培养24 h时OD600分别达到6.7和7.5;在添加645 mg/L芳樟醇时酿酒酵母BY4741已经无法生长,而ScM2和ScM6还能继续生长。上述实验证明,传代适应性驯化是改造酿酒酵母芳樟醇耐受性的有效手段。通过驯化我们成功将芳樟醇对酿酒酵母的最高致死浓度从430 mg/L提高到了645 mg/L以上。

|
| 图 5 摇瓶培养实验验证酿酒酵母突变株对芳樟醇的耐受能力 Figure 5 Detection tolerance of S. cerevisiae variant strains to linalool by liquid cultivation. A: Without addition of linalool; B: With addition of 322.5 mg/L linalool; C: With addition of 430 mg/L linalool; D: With addition of 645 mg/L linalool. A:不添加芳樟醇;B:添加322.5 mg/L芳樟醇;C:添加430 mg/L芳樟醇;D:添加645 mg/L芳樟醇 |
|
|
将ScM2、ScM6及BY4741进行了基因组重测序,并分析它们之间的差异基因。结果显示,与BY4741相比,ScM2的基因组中5个基因发生了非同义单核苷酸变异(non-synonymous single nucleotide variations,nsSNVs),ScM6基因组中4个基因发生了非同义单核苷酸位点变异。在这些突变体中,与BY4741基因组相比,ScM2和ScM6中4个基因(YBR074W、YBR172C、YHR007C和YMR275C)含有共同突变,具体如表 2所示。
| ScM2和ScM6共同或特有突变 The common or specific mutation in ScM2 and ScM6 |
突变基因 Mutated gene |
功能 Function |
突变位点 Mutational site |
| 共同突变 The common mutation |
YBR074W (pff1) | 编码多跨液泡膜蛋白酶;这是糖基化跨膜蛋白,该蛋白与金属蛋白酶M28家族同源;具有面向内腔的蛋白酶结构域,在液泡生理学中起作用 Encoding multi-spanning vacuolar membrane protease; Glycosylated transmembrane protein bearing homology to the M28 family of metalloproteases; Has a lumenal-facing protease domain; Plays role in vacuole physiology |
T838N |
| 共同突变 The common mutation |
YBR172C (smy2) | 编码GYF结构域蛋白;参与COP Ⅱ囊泡的形成;与Sec23p/Sec24p复合体相互作用 Encoding GYF domain protein; Involves in COP Ⅱ vesicle formation; Interacts with the Sec23p/Sec24p subcomplex |
K404Q |
| 共同突变 The common mutation |
YHR007C (erg11) | 编码羊毛甾醇14-α-去甲基化酶,属于细胞色素P450家族成员,在麦角甾醇生物合成途径中催化羊角甾醇C-14去甲基化形成 4, 4′-二甲基胆固-8, 14, 24-三烯-3-β-醇;麦角甾醇过量时转录下调 Encoding lanosterol 14-α-demethylase, a member of cytochrome P450 family, which catalyzes C-14 demethylation of lanosterol to form 4, 4′-dimethyl cholesta-8, 14, 24-triene-3-β-ol in ergosterol biosynthesis pathway; Transcriptionally down-regulated when ergosterol is excess |
G466R |
| 共同突变 The common mutation |
YMR275C (bul1) | 编码泛素-泛素连接酶Bul1,Rsp5p E3泛素连接酶复合物的成分;参与细胞内的多种生理过程,如:ATP输出、蛋白质泛素化等 Encoding ubiquitin-ubiquitin ligase (Bul1) that is a component of the Rsp5p E3-ubiquitin ligase complex; Involves in several processes, including ATP export and protein ubiquitination and so on |
F384C |
| ScM2特有突变 The specific mutation in ScM2 |
YLR088W (gaa1) | GPI锚定转氨酶亚基,去除GPI锚定信号并将GPI (糖基磷脂酰肌醇)连接到内质网中的蛋白质中 Subunit of the GPI: protein transamidase complex; Removes the GPI-anchoring signal and attaches GPI (glycosylphosphatidylinositol) to proteins in the endoplasmic reticulum |
A160E |
为研究上述YBR074W、YBR172C、YHR007C和YMR275C这4个基因的突变是否与酿酒酵母芳樟醇耐受性的提高有关,使用CRISPR/Cas9编辑系统对酿酒酵母BY4741基因组中的YBR074W、YBR172C、YHR007C和YMR275C基因进行了点突变,分别获得了BY4741-YBR074WT838N、BY4741-YBR172CK404Q、BY4741-YHR007CG466R和BY4741-YMR275CF384C这4株突变菌株。以野生菌株BY4741为对照组,将ScM6突变株以及BY4741-YBR074WT838N、BY4741-YBR172CK404Q、BY4741-YHR007CG466R和BY4741-YMR275CF384C菌株在含有430 mg/L芳樟醇的YPD培养基中培养,测定OD600并绘制生长曲线。结果如图 6所示,BY4741-YBR074WT838N、BY4741-YBR172CK404Q和BY4741-YHR007CG466R菌株对芳樟醇的耐受性情况与BY4741相似,在培养60 h后菌株才开始生长;BY4741-YMR275CF384C菌株与BY4741相比耐受性有所提高,菌株的生长延滞期明显缩短,24 h后菌株就开始生长,但其耐受效果明显低于ScM6菌株。猜测驯化得到的ScM6菌株耐受性提高可能是多个基因共同突变导致的结果,因此使用CRISPR/Cas9编辑系统对酿酒酵母BY4741基因组中的YBR074W、YBR172C、YHR007C和YMR275C基因进行同时点突变,获得了BY4741-M4菌株;将其在添加430 mg/L芳樟醇的YPD培养基中进行培养,结果显示BY4741-M4菌株的生长情况与ScM6菌株相近(图 6)。上述结果说明YBR074WT838N、YBR172CK404Q、YHR007CG466R和YMR275CF384C这4个基因突变对耐受性的提高具有重要作用。

|
| 图 6 添加430 mg/L芳樟醇时突变菌株的生长曲线 Figure 6 Growth curves of S. cerevisiae mutants with addition of 430 mg/L of linalool. |
|
|
为了测试进化菌株的遗传稳定性,对ScM2和ScM6菌株进行了传代实验。在YPD培养基中添加了430 mg/L芳樟醇,比较了第1、10、15和20代菌株的生长情况。结果如图 7所示,在进行了连续20次传代后,ScM2和ScM6菌株生长较1代无明显变化,而与野生型BY4741菌株相比仍表现出了明显的生长优势。说明ScM2和ScM6菌株的稳定性较好,在连续培养过程中仍维持了较好的芳樟醇耐受性能。

|
| 图 7 ScM2和ScM6菌株遗传稳定性分析培养基中添加了430 mg/L芳樟醇 Figure 7 The genetic stability analysis of ScM2 and ScM6. The culture medium was added 430 mg/L linalool. |
|
|
酿酒酵母作为基因操作简单的真核微生物,在生物安全性方面和植物基因的功能表达方面有其独特的优势,是植物天然产物异源合成的极佳宿主。近年来,研究者通过蛋白质工程和代谢工程改造手段已经实现了多种萜类化合物(如青蒿素、番茄红素等)在酿酒酵母中的高效合成。但是单萜的酿酒酵母合成却相对落后,其中单萜对微生物毒性强是限制其高效合成的一个关键因素。在本研究中,根据酵母菌在不同浓度芳樟醇处理下的生长曲线,发现芳樟醇的浓度为172 mg/L时酿酒酵母的生长已经受到轻微的影响,而当芳樟醇的浓度达到430 mg/L时酿酒酵母的生长受到了抑制。虽然国内外研究者目前已逐步揭示了单萜对微生物的一些毒害因素以及微生物在单萜胁迫下的应答机制[16, 18-19, 29],但有关酿酒酵母芳樟醇耐受性改造方面的研究较少。由于单萜会诱发细胞内ROS的积累并破坏细胞的细胞膜,从而影响菌体正常生长。因此,为了提高酿酒酵母对芳樟醇的耐受性,本研究首先从减少胞内毒性物质的积累、清除胞内的ROS及强化细胞壁膜功能这几个角度对酿酒酵母芳樟醇耐受性进行改造。
ABC转运蛋白是一类超大膜转运蛋白,可以通过ATP驱动将无机盐、氨基酸、多肽、烷烃等物质进行逆浓度梯度跨膜运输。多向耐药性(pleiotropic drug resistance,PDR)蛋白和多药耐药性相关蛋白(multidrug resistance-associated protein,MRP)是ABC转运蛋白的2个亚族,可以识别和转运不同类型的底物,包括抗生素、黄酮类、萜类等化学品。有研究表明,在烷烃胁迫下,PDR蛋白Snq2、Pdr5、Pdr15及MRP蛋白Yor1显著上调[21]。Ling等发现过表达外排泵Snq2和Pdr5可以降低细胞内C10和C11烷烃化合物的水平,从而提高对C10和C11烷烃化合物的耐受性[21]。此外,有研究者利用强启动子上调Pdr18外排蛋白的表达,使得酿酒酵母乙醇耐受性和产量都明显提高[30]。然而在本研究中,分别过表达Yor1、Snq2、Pdr5、Pdr15和Pdr18这5个ABC转运蛋白并不能提高酿酒酵母对芳樟醇的耐受性。类似地,研究者在酿酒酵母对D-柠檬烯耐受性的实验中发现,pdr5、yor1和pdr15转录水平虽然上调,但过表达这些蛋白也不能提高酿酒酵母对D-柠檬烯(C10烯烃类物质)的耐受性[31]。我们猜测特定物质需要与特定转运蛋白结合以及物质转运效率与蛋白质结构有着密切的关系,为此今后的研究中可以筛选特定的芳樟醇转运蛋白或者对有转运功能的蛋白进行分子改造,从而提高转运效率。此外,虽然过表达ABC转运蛋白可以促进产物外排,减少毒性物质在细胞内的积累,但ABC转运蛋白作为跨膜蛋白,过量表达也会造成细胞的代谢负担。
一些研究表明单萜会诱发细胞内活性氧的积累[18],活性氧会破坏细胞中的脂质、蛋白质、碳水化合物和核酸,从而造成细胞毒性。在D-柠檬烯作用下,谷氧还蛋白系统相关的基因gpx2、gsh2和硫氧还蛋白系统相关基因trr1会上调表达,而硫氧还蛋白系统相关基因tsa1和ahp1会下调表达,从而进行自由基解毒[18]。另外,其他应激响应相关基因如gre1、gre2、ctt1、hsp12和yhb1等也会被诱导表达来调控ROS水平[21]。此外,在应对ROS等有害物质时,Yap1、Yap5、Stb5和Ino2等这些转录调控因子会调控抗氧化相关基因的转录。其中Ino2调控因子除了调控自由基相关基因外,可以同时调控与外排泵、脂质代谢和麦角固醇合成相关基因的表达[21]。在本研究中,我们利用强启动子过表达了ROS调控相关的酶(Gre2、Ctt1、Yhb1、Gpx2、Trr1、Trx2和Gsh2)及4个转录调控因子(Ino2、Yap1、Yap5、Stb5),但在酿酒酵母对芳樟醇的耐受性测试中,这些基因的单独过表达也并未有效改善酿酒酵母对芳樟醇的耐受情况。酿酒酵母抵御氧胁迫的调控网络复杂,通常存在由小分子化合物如谷胱甘肽组成的非酶保护系统和抗氧化酶保护系统,用于清除细胞内的ROS,维持细胞氧化还原平衡。目前对于芳樟醇引发的毒性机理仍不清楚,菌株抗逆的分子机制通常比较复杂,很难通过单一的遗传因素改造提高菌株耐受性。为了提高酿酒酵母芳樟醇的耐受性,可能需要进行多方面的协调改造。
在本研究中,我们进一步采取了传代适应性驯化方法,通过酿酒酵母基因组自然突变及芳樟醇压力选择性筛选,获得两株耐受性提高的酿酒酵母突变株。通过全基因重测序分析,发现这两株突变株都含有YBR074WT838N、YBR172CK404Q、YHR007CG466R、YMR275CF384C这4个基因突变。在进行耐受性实验验证时,发现对酿酒酵母进行单个基因突变时,菌株耐受性不比驯化筛选出来的菌株强,但当对4个基因进行同时突变时,菌株表现出了与驯化菌株一样的耐受性能。目前尚无关于这4个基因与菌株耐受性能相关的文献报道,但对4个基因的功能有一定的研究。YBR074W编码的是一个糖基化的多跨液泡膜蛋白酶(Pff1),它的蛋白酶结构域面向液泡内腔,与液泡的运输和渗透压调节等密切相关[32]。YBR172C编码的是一个GYF结构域蛋白,该蛋白与外壳蛋白复合物Ⅱ (COPⅡ)囊泡的形成相关,会与内质网到高尔基体囊泡运输相关基因相互作用[33]。YHR007C编码羊毛甾醇14-α-去甲基化酶(Erg11),该酶是一个P450酶,在麦角甾醇生物合成途径中催化羊角甾醇C-14去甲基化形成4, 4′-二甲基胆固醇-8, 14, 24-三烯-3-β-醇,是细胞膜中麦角甾醇生物合成中的一个关键酶[34]。YMR275C编码泛素-泛素连接酶(ubiquitin-ubiquitin ligase) Bul1,Rsp5p E3泛素连接酶复合物的泛素结合成分,属于泛素化辅助蛋白;参与细胞内的多种生理过程,如ATP输出、蛋白质泛素化等[35-36]。敲除Bul1会导致细胞生长对温度敏感[35],过表达Bul1会使Gap1蛋白(广谱氨基酸透性酶,是一种通用型的氨基酸运输载体)被转运至液泡中,从而导致其被降解[36]。目前关于YBR074WT838N、YBR172CK404Q、YHR007CG466R和YMR275CF384C这4个基因突变对耐受性提高的具体机制还不清楚。在今后的研究中,可以通过这4个基因的过表达或者下调、敲除,进一步探讨4个基因对耐受性的影响机制。此外,可进一步对驯化菌株进行转录组学分析,研究耐受性提高菌株在基因转录水平上的变化,阐明耐受性提高的具体机制。
传代适应性驯化不需要考虑复杂的代谢调控网络,只需要利用特定的胁迫压力就可以富集并筛选出耐受性能提高的目标突变株,这种方法简单、高效、适用性广[37],很多研究者利用这种方法提高了菌株对甲醇[38]、柠檬烯[24]、温度[39]等的耐受性能。然而传代适应性驯化过程耗时较长,基因组的随机多重突变使后续耐受性的机制解析变得复杂。近年来,随着基因工程技术的发展,研究者开发出了多种基因组进化技术策略,如转座子插入诱变、RNAi辅助基因组进化方法、基于CRISPR/Cas的可追踪基因组进化方法等[40],其中基于CRISPR系统的基因组进化技术速度快且针对性强、突变范围广,弥补了传代适应进化技术的一些缺陷,有利于菌株耐受性能的改良和机制解析。如果能结合运用多种基因组进化技术,今后将更大程度地提高酿酒酵母对芳樟醇等单萜的耐受性能。
本研究中,通过单独过表达ABC转运蛋白(Yor1、Snq2、Pdr5、Pdr15和Pdr18)、ROS调控相关酶(Gre2、Ctt1、Yhb1、Gpx2、Trr1、Trx2和Gsh2)及转录调控因子(Ino2、Yap1、Yap5和Stb5)未能有效提高酿酒酵母芳樟醇的耐受性。然而通过对酵母细胞进行传代适应性驯化,获得了两株芳樟醇耐受性能力提高的酿酒酵母突变菌株,在液体培养中,将芳樟醇对酿酒酵母的最大致死浓度从430 mg/L提高到了645 mg/L以上。通过基因组重测序分析,获得了4个发生单核苷酸突变的基因,其中表达YMR275CF384C突变基因一定程度上可以提高酵母芳樟醇耐受性,但耐受性效果明显低于驯化中筛选的ScM6菌株。同时对YBR074WT838N、YBR172CK404Q、YHR007CG466R和YMR275CF384C这4个基因进行突变后,菌株的耐受性明显提高。本研究为芳樟醇和其他单萜的耐受性改造提供了参考。
| [1] |
Karuppiah V, Ranaghan KE, Leferink NGH, Johannissen LO, Shanmugam M, Ní Cheallaigh A, Bennett NJ, Kearsey LJ, Takano E, Gardiner JM, et al. Structural basis of catalysis in the bacterial monoterpene synthases linalool synthase and 1, 8-cineole synthase[J]. ACS Catalysis, 2017, 7(9): 6268-6282. DOI:10.1021/acscatal.7b01924 |
| [2] |
Aprotosoaie AC, Hăncianu M, Costache Ⅱ, Miron A. Linalool: a review on a key odorant molecule with valuable biological properties[J]. Flavour and Fragrance Journal, 2014, 29(4): 193-219. DOI:10.1002/ffj.3197 |
| [3] |
Lapczynski A, Letizia CS, Api AM. Addendum to fragrance material review on linalool[J]. Food and Chemical Toxicology, 2008, 46(11): S190-S192. DOI:10.1016/j.fct.2008.06.087 |
| [4] |
Marienhagen J, Bott M. Metabolic engineering of microorganisms for the synthesis of plant natural products[J]. Journal of Biotechnology, 2013, 163(2): 166-178. DOI:10.1016/j.jbiotec.2012.06.001 |
| [5] |
Deng Y, Sun MX, Xu S, Zhou JW. Enhanced (S)-linalool production by fusion expression of farnesyl diphosphate synthase and linalool synthase in Saccharomyces cerevisiae[J]. Journal of Applied Microbiology, 2016, 121(1): 187-195. DOI:10.1111/jam.13105 |
| [6] |
Luo YZ, Li BZ, Liu D, Zhang L, Chen Y, Jia B, Zeng BX, Zhao HM, Yuan YJ. Engineered biosynthesis of natural products in heterologous hosts[J]. Chemical Society Reviews, 2015, 44(15): 5265-5290. DOI:10.1039/C5CS00025D |
| [7] |
Zebec Z, Wilkes J, Jervis AJ, Scrutton NS, Takano E, Breitling R. Towards synthesis of monoterpenes and derivatives using synthetic biology[J]. Current Opinion in Chemical Biology, 2016, 34: 37-43. DOI:10.1016/j.cbpa.2016.06.002 |
| [8] |
Sun MX, Liu JD, Du GC, Zhou JW, Chen J. Regulation of isoprenoid pathway for enhanced production of linalool in Saccharomyces cerevisiae[J]. Chinese Journal of Biotechnology, 2013, 29(6): 751-759. (in Chinese) 孙明雪, 刘继栋, 堵国成, 周景文, 陈坚. 调控酿酒酵母类异戊二烯合成途径强化芳樟醇合成[J]. 生物工程学报, 2013, 29(6): 751-759. |
| [9] |
Fu BB, Zhao JZ, Li C, Liu XL, Bao XM, Hou J. Research progresses on monoterpene synthesis in Saccharomyces cerevisiae[J]. Biotechnology Bulletin, 2018, 34(4): 60-69. (in Chinese) 伏贝贝, 赵建志, 李琛, 刘新利, 鲍晓明, 侯进. 酿酒酵母单萜合成的研究进展[J]. 生物技术通报, 2018, 34(4): 60-69. |
| [10] |
Amiri P, Shahpiri A, Asadollahi MA, Momenbeik F, Partow S. Metabolic engineering of Saccharomyces cerevisiae for linalool production[J]. Biotechnology Letters, 2016, 38(3): 503-508. DOI:10.1007/s10529-015-2000-4 |
| [11] |
Cao X, Wei LJ, Lin JY, Hua Q. Enhancing linalool production by engineering oleaginous yeast Yarrowia lipolytica[J]. Bioresource Technology, 2017, 245: 1641-1644. DOI:10.1016/j.biortech.2017.06.105 |
| [12] |
Peng BY, Nielsen LK, Kampranis SC, Vickers CE. Engineered protein degradation of farnesyl pyrophosphate synthase is an effective regulatory mechanism to increase monoterpene production in Saccharomyces cerevisiae[J]. Metabolic Engineering, 2018, 47: 83-93. DOI:10.1016/j.ymben.2018.02.005 |
| [13] |
Zhou PP, Du Y, Xu NN, Yue CL, Ye LD. Improved linalool production in Saccharomyces cerevisiae by combining directed evolution of linalool synthase and overexpression of the complete mevalonate pathway[J]. Biochemical Engineering Journal, 2020, 161: 107655. DOI:10.1016/j.bej.2020.107655 |
| [14] |
Zhou PP, Du Y, Fang X, Xu NN, Yue CL, Ye LD. Combinatorial modulation of linalool synthase and farnesyl diphosphate synthase for linalool overproduction in Saccharomyces cerevisiae[J]. Journal of Agricultural and Food Chemistry, 2021, 69(3): 1003-1010. DOI:10.1021/acs.jafc.0c06384 |
| [15] |
Liu J, Zhu Y, Du G, Zhou J, Chen J. Exogenous ergosterol protects Saccharomyces cerevisiae from D-limonene stress[J]. Journal of Applied Microbiology, 2013, 114(2): 482-491. DOI:10.1111/jam.12046 |
| [16] |
Brennan TCR, Krömer JO, Nielsen LK. Physiological and transcriptional responses of Saccharomyces cerevisiae to D-limonene show changes to the cell wall but not to the plasma membrane[J]. Applied and Environmental Microbiology, 2013, 79(12): 3590-3600. DOI:10.1128/AEM.00463-13 |
| [17] |
Bakkali F, Averbeck S, Averbeck D, Zhiri A, Idaomar M. Cytotoxicity and gene induction by some essential oils in the yeast Saccharomyces cerevisiae[J]. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 2005, 585(1/2): 1-13. |
| [18] |
Liu JD, Zhu YB, Du GC, Zhou JW, Chen J. Response of Saccharomyces cerevisiae to D-limonene-induced oxidative stress[J]. Applied Microbiology and Biotechnology, 2013, 97(14): 6467-6475. DOI:10.1007/s00253-013-4931-9 |
| [19] |
Uribe S, Ramirez J, Peña A. Effects of beta-pinene on yeast membrane functions[J]. Journal of Bacteriology, 1985, 161(3): 1195-1200. DOI:10.1128/jb.161.3.1195-1200.1985 |
| [20] |
Zhang L, Xiao WH, Wang Y, Yao MD, Jiang GZ, Zeng BX, Zhang RS, Yuan YJ. Chassis and key enzymes engineering for monoterpenes production[J]. Biotechnology Advances, 2017, 35(8): 1022-1031. DOI:10.1016/j.biotechadv.2017.09.002 |
| [21] |
Ling H, Chen BB, Kang A, Lee JM, Chang MW. Transcriptome response to alkane biofuels in Saccharomyces cerevisiae: identification of efflux pumps involved in alkane tolerance[J]. Biotechnology for Biofuels, 2013, 6(1): 95. DOI:10.1186/1754-6834-6-95 |
| [22] |
Li J, Zhu K, Miao L, Rong LX, Zhao Y, Li SL, Ma LJ, Li JX, Zhang CY, Xiao DG, et al. Simultaneous improvement of limonene production and tolerance in Yarrowia lipolytica through tolerance engineering and evolutionary engineering[J]. ACS Synthetic Biology, 2021, 10(4): 884-896. DOI:10.1021/acssynbio.1c00052 |
| [23] |
Wang Y, Lim L, DiGuistini S, Robertson G, Bohlmann J, Breuil C. A specialized ABC efflux transporter GcABC-G1 confers monoterpene resistance to Grosmannia clavigera, a bark beetle-associated fungal pathogen of pine trees[J]. New Phytologist, 2013, 197(3): 886-898. DOI:10.1111/nph.12063 |
| [24] |
Brennan TCR, Williams TC, Schulz BL, Palfreyman RW, Krömer JO, Nielsen LK. Evolutionary engineering improves tolerance for replacement jet fuels in Saccharomyces cerevisiae[J]. Applied and Environmental Microbiology, 2015, 81(10): 3316-3325. DOI:10.1128/AEM.04144-14 |
| [25] |
Xiao L, Wu AN, Xu GC, Han RZ, Ni Y. Adapting n-butanol tolerance of Pseudomonas putida and comparative genomics analysis[J]. Journal of Food Science and Biotechnology, 2020, 39(10): 65-72. (in Chinese) 肖琳, 吴安宁, 许国超, 韩瑞枝, 倪晔. 恶臭假单胞菌的正丁醇耐受性驯化及比较基因组学分析[J]. 食品与生物技术学报, 2020, 39(10): 65-72. DOI:10.3969/j.issn.1673-1689.2020.10.009 |
| [26] |
Zhou PP, Ye LD, Xie WP, Lv XM, Yu HW. Highly efficient biosynthesis of astaxanthin in Saccharomyces cerevisiae by integration and tuning of algal crtZ and bkt[J]. Applied Microbiology and Biotechnology, 2015, 99(20): 8419-8428. DOI:10.1007/s00253-015-6791-y |
| [27] |
Gietz RD, Schiestl RH. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method[J]. Nature Protocols, 2007, 2(1): 31-34. DOI:10.1038/nprot.2007.13 |
| [28] |
Zhou PP, Fang X, Xu NN, Yao Z, Xie WP, Ye LD. Development of a highly efficient copper-inducible GAL regulation system (CuIGR) in Saccharomyces cerevisiae[J]. ACS Synthetic Biology, 2021, 10(12): 3435-3444. DOI:10.1021/acssynbio.1c00378 |
| [29] |
Parveen M, Hasan MK, Takahashi J, Murata Y, Kitagawa E, Kodama O, Iwahashi H. Response of Saccharomyces cerevisiae to a monoterpene: evaluation of antifungal potential by DNA microarray analysis[J]. Journal of Antimicrobial Chemotherapy, 2004, 54(1): 46-55. DOI:10.1093/jac/dkh245 |
| [30] |
Teixeira MC, Godinho CP, Cabrito TR, Mira NP, Sá-Correia I. Increased expression of the yeast multidrug resistance ABC transporter Pdr18 leads to increased ethanol tolerance and ethanol production in high gravity alcoholic fermentation[J]. Microbial Cell Factories, 2012, 11: 98. DOI:10.1186/1475-2859-11-98 |
| [31] |
Hu FF, Liu JD, Du GC, Hua ZZ, Zhou JW, Chen J. Key cytomembrane ABC transporters of Saccharomyces cerevisiae fail to improve the tolerance to D-limonene[J]. Biotechnology Letters, 2012, 34(8): 1505-1509. DOI:10.1007/s10529-012-0931-6 |
| [32] |
Hecht KA, Wytiaz VA, Ast T, Schuldiner M, Brodsky JL. Characterization of an M28 metalloprotease family member residing in the yeast vacuole[J]. FEMS Yeast Research, 2013, 13(5): 471-484. DOI:10.1111/1567-1364.12050 |
| [33] |
Higashio H, Sato K, Nakano A. Smy2p participates in COPⅡ vesicle formation through the interaction with Sec23p/Sec24p subcomplex[J]. Traffic, 2008, 9(1): 79-93. DOI:10.1111/j.1600-0854.2007.00668.x |
| [34] |
Turi TG, Loper JC. Multiple regulatory elements control expression of the gene encoding the Saccharomyces cerevisiae cytochrome P450, lanosterol 14 alpha-demethylase (ERG11)[J]. Journal of Biological Chemistry, 1992, 267(3): 2046-2056. DOI:10.1016/S0021-9258(18)46051-6 |
| [35] |
Yashiroda H, Oguchi T, Yasuda Y, Toh-E A, Kikuchi Y. Bul1, a new protein that binds to the Rsp5 ubiquitin ligase in Saccharomyces cerevisiae[J]. Molecular and Cellular Biology, 1996, 16(7): 3255-3263. DOI:10.1128/MCB.16.7.3255 |
| [36] |
Merhi A, André B. Internal amino acids promote Gap1 permease ubiquitylation via TORC1/Npr1/14-3-3-dependent control of the Bul arrestin-like adaptors[J]. Molecular and Cellular Biology, 2012, 32(22): 4510-4522. DOI:10.1128/MCB.00463-12 |
| [37] |
Sandberg TE, Salazar MJ, Weng LL, Palsson BO, Feist AM. The emergence of adaptive laboratory evolution as an efficient tool for biological discovery and industrial biotechnology[J]. Metabolic Engineering, 2019, 56: 1-16. |
| [38] |
Wang Y, Fan LW, Tuyishime P, Liu J, Zhang K, Gao N, Zhang ZH, Ni XM, Feng JH, Yuan QQ, et al. Adaptive laboratory evolution enhances methanol tolerance and conversion in engineered Corynebacterium glutamicum[J]. Communications Biology, 2020, 3: 217. DOI:10.1038/s42003-020-0954-9 |
| [39] |
Caspeta L, Chen Y, Ghiaci P, Feizi A, Buskov S, Hallström BM, Petranovic D, Nielsen J. Altered sterol composition renders yeast thermotolerant[J]. Science, 2014, 346(6205): 75-78. DOI:10.1126/science.1258137 |
| [40] |
Xia SY, Jiang LH, Cai J, Huang L, Xu ZN, Lian JZ. Advances in genome evolution of Saccharomyces cerevisiae[J]. Synthetic Biology Journal, 2020, 1(5): 556-569. (in Chinese) 夏思杨, 江丽红, 蔡谨, 黄磊, 徐志南, 连佳长. 酿酒酵母基因组进化的研究进展[J]. 合成生物学, 2020, 1(5): 556-569. |