 2021, Vol. 48
2021, Vol. 48扩展功能
文章信息
- 王慧慧, 张非, 舒潼, 李攀登, 吴亚, 余天意, 付春华, 余龙江
- WANG Huihui, ZHANG Fei, SHU Tong, LI Pandeng, WU Ya, YU Tianyi, FU Chunhua, YU Longjiang
- 两株芽孢杆菌在苎麻纤维复合脱胶中的应用
- Application of two Bacillus sp. strains in composite degumming of ramie fibers
- 微生物学通报, 2021, 48(8): 2512-2523
- Microbiology China, 2021, 48(8): 2512-2523
- DOI: 10.13344/j.microbiol.china.201092
-
文章历史
- 收稿日期: 2020-11-21
- 接受日期: 2021-02-02
- 网络首发日期: 2021-03-16





苎麻纤维细长、透气性好、传热快、散湿快、纤维强力大且轻盈、不易霉变,被誉为“天然纤维之王”,广泛用于纺织、医疗、军工等行业[1-2]。但苎麻韧皮纤维中含有30%左右的胶质,主要包括半纤维素、果胶等[3-4]。胶质的存在会影响苎麻纤维的质量,必须把这些胶质去除,以得到精干麻,即脱胶[5-6]。传统的脱胶工艺使用大量的酸碱,不仅污染环境,还会损伤麻纤维,导致精干麻品质差[7-8];酶脱胶一般使用商品酶,成本较高且效率低,无法实现大规模工业生产[9-10];而微生物脱胶条件温和,绿色环保,越来越受到学者的关注[11-13]。
目前报道的应用于苎麻脱胶的微生物较多,主要集中在芽孢杆菌属、假单胞菌属、肠杆菌属、欧文氏菌属和曲霉菌等[14-16],其特征主要是能产生降解苎麻胶质的酶类,如果胶酶、半纤维素酶。然而,多数菌株分泌的酶类只有一种或几种,或者缺少脱胶关键酶,导致脱胶处理达不到麻纺对纤维残胶率的要求[17]。复合微生物脱胶是近年来研究的热点,通常是将2株或以上的菌株同时或者依次作用于原麻,不同微生物所利用的胶质成分、产酶顺序、分泌的酶类组成比例及水解方式相互协同,从而逐步实现胶质成分的有效降解,提高脱胶效果。然而,苎麻复合微生物脱胶虽然取得一定进展,但仍存在脱胶体系中关键酶系不足,脱胶时间长达几十小时,脱胶效率比较低,这些限制了苎麻生物脱胶的大规模生产[18-19]。因此,发掘合适的微生物进行复合脱胶及工艺优化仍是苎麻生物脱胶技术发展的关键。
本研究以实验室保藏的高产果胶酶、甘露聚糖酶的芽孢杆菌HG-9和高产木聚糖酶的芽孢杆菌HG-25为基础,进行混合脱胶,并进一步对复合微生物脱胶条件进行优化,不仅促进了苎麻胶质的进一步去除,缩短了脱胶时间,而且减少了脱胶后期漂白剂的用量,具有良好的经济效益和环境效益。
1 材料与方法 1.1 苎麻韧皮苎麻韧皮由湖北精华纺织集团有限公司提供,并于干燥阴凉处保藏。
1.2 菌株芽孢杆菌HG-9 (CCTCC号:M2014439)和芽孢杆菌HG-25 (CCTCC号:M2014438)由本实验室筛选保藏。
1.3 主要试剂、仪器和培养基酵母提取物、胰蛋白胨,安琪酵母股份有限公司;氯化钠,国药集团化学试剂有限公司;琼脂粉,Biosharp Life Sciences公司。
恒温摇床、恒温培养箱,武汉瑞华仪器设备有限责任公司;超净工作台,苏州安泰空气技术有限公司;电子显微镜,FEI公司;pH计、电子分析天平,梅特勒托利多仪器(上海)有限公司;电热鼓风干燥箱,天津市泰斯特仪器有限公司;立式压力蒸汽灭菌器,上海博迅医疗生物仪器股份有限公司。
LB液体培养基(g/L):酵母提取物5.0,胰蛋白胨10.0,NaCl 10.0,pH 7.0,1×105 Pa灭菌20 min。LB固体培养基(g/L):酵母提取物5.0,胰蛋白胨10.0,NaCl 10.0,琼脂粉15.0,pH 7.0,1×105 Pa灭菌20 min。
1.4 菌株HG-9和HG-25的扩大培养取保藏的菌株HG-9和HG-25,划线法涂布于装有LB固体培养基的培养皿,37 ℃倒置培养。然后挑取单菌落至LB液体培养基,37 ℃、180 r/min分别培养8 h和10 h。
1.5 菌株HG-9和HG-25单独脱胶研究 1.5.1 菌株HG-9单独脱胶研究取5 g苎麻,置于装有60 mL水的250 mL锥形瓶中,接入5 mL培养8 h (此时菌体生长最为旺盛,达到3.0×109 CFU/mL)的菌株HG-9菌悬液。脱胶初始pH自然,37 ℃、180 r/min脱胶16 h。脱胶过程中每隔2 h取2 mL脱胶液,采用二硝基水杨酸(DNS)法[20]测定其果胶酶活力、甘露聚糖酶活力、木聚糖酶活力和纤维素酶活力。
果胶酶活力测定原理:果胶酶作用下,果胶可被降解为还原性糖,DNS在碱性条件下可与还原糖发生反应,产物在沸水浴条件下显色。产物颜色的深浅与生成的还原糖的量成正比,从而通过反应液颜色表征脱胶液中果胶酶的活力。
果胶酶活测定:取1.8 mL果胶溶液于试管中,加入0.2 mL脱胶上清液,37 ℃反应30 min,然后加入3 mL DNS溶液混合均匀置于沸水浴条件下5 min,冰水冷却后测定OD540吸光值。
酶活性定义:在特定条件(37 ℃、pH 7.0)下,每分钟内转化生成1 μmol半乳糖醛酸所需酶量为一个酶活性单位U。
甘露聚糖酶、木聚糖酶和纤维素酶活力测定时底物分别为甘露聚糖、木聚糖和羧甲基纤维素钠,产物分别为甘露糖、木糖和葡萄糖。
脱胶完成后,用自来水将苎麻样品冲洗干净,置于称量瓶中,105 ℃烘至恒重后于电子分析天平称重,并对菌株HG-9处理前后苎麻组成成分进行分析。
1.5.2 菌株HG-25单独脱胶研究取5 g苎麻,置于装有60 mL水的250 mL锥形瓶中,接入4.8 mL培养10 h (此时菌体生长最为旺盛,达到2.5×109 CFU/mL)的菌株HG-25菌悬液。脱胶初始pH为6.0,于35 ℃、180 r/min脱胶16 h。脱胶过程中每隔2 h取2 mL脱胶液,分别测定果胶酶活力、甘露聚糖酶活力、木聚糖酶活力和纤维素酶活力。
酶活测定方法同1.5.1。
脱胶完成后,用自来水将苎麻样品冲洗干净,置于称量瓶中,105 ℃烘至恒重后于电子分析天平称重,并对菌株HG-25处理前后苎麻组成成分进行分析。
1.6 菌株HG-9/HG-25混合脱胶研究 1.6.1 混合菌接种量的正交优化根据菌株HG-9和HG-25单独脱胶特点,采用L16 (42)正交设计,以脱胶率为指标确定2株菌株最佳接种比例,因素水平见表 1。通过正交试验获得最佳接种量组合,并对该条件下的苎麻脱胶率进行测定。
| 水平/因素 Level/Factor |
菌株HG-9 (A)接种量 Inoculum size of strain HG-9 (A) (%) |
菌株HG-25 (B)接种量 Inoculum size of strain HG-25 (B) (%) |
| 1 | 4 | 4 |
| 2 | 6 | 6 |
| 3 | 8 | 8 |
| 4 | 10 | 10 |
根据菌株HG-9和HG-25单独脱胶条件,采用Box-Behnken Design响应面法对苎麻混合菌脱胶条件进行优化。以苎麻脱胶率作为响应值,设计三因素三水平的响应面分析法,因素水平见表 2。对优化条件下的脱胶率及胶质去除率进行测定。
| 水平 Level |
因素 Factor |
||
| 水料比 Water-to-material ratio |
初始pH值 Initial pH |
脱胶温度 Degumming temperature (℃) |
|
| -1 | 12:1 | 5 | 32 |
| 0 | 14:1 | 6 | 37 |
| 1 | 16:1 | 7 | 42 |
取5 g苎麻样品置于装有80 mL水的锥形瓶中,接种培养8 h的HG-9菌悬液和培养10 h的HG-25菌悬液,接种量均为6%,脱胶初始pH为5.9,在37.6 ℃和180 r/min下分别处理12、14、16、18、20 h,并对苎麻脱胶率及相应的胶质去除率进行分析。
1.7 漂白剂使用量研究取5 g苎麻样品,菌株HG-9单独脱胶及HG-9/HG-25混合脱胶后,加入0.25 g氢氧化钠和100 mL水,在冷凝回流条件下煮沸2 h。取出苎麻,在流水下冲洗干净,置于装有100 mL水的烧杯中,分别加入0.06、0.08、0.10、0.12、0.14 g次氯酸钠,漂白5 min,对漂白后的苎麻进行洗涤、干燥,并测定苎麻残胶率。
1.8 苎麻各组成成分含量、脱胶率和残胶率的测定苎麻各组成成分含量、含胶率及残胶率测定根据国家标准GB/T 5889-1986《苎麻化学成分定量分析方法》[21]。

|
公式中M1为苎麻原麻恒重质量(g);M2为苎麻原麻脱胶后恒重质量(g)。
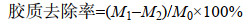
|
公式中M0为苎麻原麻的胶质含量(g);M1为苎麻原麻恒重质量(g);M2为苎麻原麻脱胶后恒重质量(g),本研究中所用苎麻韧皮胶质含量为27.99%。
1.9 苎麻纤维微观形态分析将苎麻原麻及不同微生物处理后的苎麻样品剪切至约0.50 cm后制样,于电子显微镜下观察其微观形态差异。该分析由华中科技大学分析测试中心测定。
2 结果与讨论 2.1 菌株HG-9苎麻脱胶研究由图 1可知,利用菌株HG-9单独脱胶时,其产生的脱胶酶系主要包括果胶酶和甘露聚糖酶,而且随着脱胶过程的进行,它们的酶活力逐渐增加,在8 h时分别达到118.02 U/mL和99.48 U/mL。此外,其产生的木聚糖酶活性较低,在8 h时仅为4.88 U/mL,而且其基本不产纤维素酶,从而避免了对苎麻纤维素的损伤。

|
| 图 1 菌株HG-9在苎麻脱胶过程中各酶活变化趋势 Figure 1 The change trend of enzyme activity during ramie degumming of strain HG-9 |
|
|
菌株HG-9脱胶处理16 h后苎麻各组成成分变化如图 2所示,可以看出其脱胶率可达12.2%,胶质去除率达43.54%。其中,果胶和水溶物去除效果较好,含量从5.32%和5.75%分别降至0.75%和0.86%,去除率可达85.90%和85.04%;而半纤维素去除效果较差,其含量从13.89%降至11.83%,去除率只有14.83%,可见,甘露聚糖酶不足以去除苎麻半纤维素,推测木聚糖酶也是苎麻脱胶的关键酶。此外,其木质素含量从1.45%降至1.30%,去除率只有10.34%。在结构上,木质素通过共价键与半纤维素连接,推测半纤维素的去除可带动木质素的去除。基于对菌株HG-9脱胶能力的分析,选择一株具有高木聚糖酶活性的菌株以实现二者的高效协同脱胶十分重要。

|
| 图 2 菌株HG-9脱胶处理前后苎麻化学成分分析 Figure 2 Analysis of chemical components of ramie before and after degumming by strain HG-9 |
|
|
为研究菌株HG-25的脱胶性能,对其脱胶过程中的关键酶活力进行了测定(图 3)。由图 3可知,菌株HG-25在单独脱胶时所产酶系主要包括木聚糖酶和甘露聚糖酶,而且随着脱胶过程的进行,它们的酶活力逐渐增加,在10 h时木聚糖酶活性到达最大值,为44.83 U/ml,甘露聚糖酶在8 h时活力最高,为48.25 U/ml;而其产生的果胶酶活性较低,而且不产纤维素酶,从而避免了对苎麻纤维素的损伤。

|
| 图 3 菌株HG-25在苎麻脱胶过程中各酶活变化趋势 Figure 3 The change trend of enzyme activity during ramie degumming of strain HG-25 |
|
|
菌株HG-25脱胶处理苎麻16 h后各成分变化见图 4。可以看到,经菌株HG-25处理后的苎麻其半纤维素含量由13.89%降至9.15%,去除率达34.12%。菌株HG-25在产酶和胶质去除上均与HG-9具备很好的互补性。因此,基于HG-9的单菌种脱胶中缺少脱胶关键酶木聚糖酶引起脱胶不完全、纤维品质低的问题,菌株HG-25的引入对进一步提高苎麻脱胶率,特别是半纤维素去除率具有重要作用。

|
| 图 4 菌株HG-25脱胶处理前后苎麻化学成分分析 Figure 4 Analysis of chemical components of ramie before and after degumming by strain HG-25 |
|
|
以苎麻脱胶率为指标,采用L16 (42)正交设计对菌株HG-9和HG-25混合脱胶的最佳接种量进行优化(表 3)。
| 实验编号 Experiment No. |
A HG-9接种量 A Inoculum size of HG-9 |
B HG-25接种量 B Inoculum size of HG-25 |
脱胶率 Degumming rate (%) |
| 1 | 1 | 2 | 13.40 |
| 2 | 2 | 2 | 14.21 |
| 3 | 4 | 1 | 12.81 |
| 4 | 4 | 2 | 12.59 |
| 5 | 3 | 1 | 12.78 |
| 6 | 4 | 3 | 11.90 |
| 7 | 2 | 4 | 13.36 |
| 8 | 2 | 3 | 14.29 |
| 9 | 2 | 1 | 14.20 |
| 10 | 4 | 4 | 11.24 |
| 11 | 3 | 4 | 12.04 |
| 12 | 3 | 3 | 13.95 |
| 13 | 3 | 2 | 14.31 |
| 14 | 1 | 3 | 11.87 |
| 15 | 1 | 4 | 11.29 |
| 16 | 1 | 1 | 13.77 |
| K1 | 13.20 | 13.39 | |
| K2 | 14.02 | 13.63 | |
| K3 | 13.27 | 13.00 | |
| K4 | 12.14 | 11.98 | |
| R | 1.88 | 1.65 |
表 3中A代表菌株HG-9接种量,B代表菌株HG-25接种量,1-4分别代表接种量4%、6%、8%和10%。根据K值大小可知菌株HG-9和HG-25接种量的最佳组合为A2B2,即接种量均为6%。在该优化条件下进行脱胶试验验证,苎麻脱胶率可达14.30%±0.13%,提高了0.27%,可用于二者混合脱胶。在微生物混合脱胶过程中,接种量过低不利于菌体的生长增殖,接种量过高,又会给生长环境带来压力,因此,优化合适的接种量十分有必要。
2.4 微生物混合脱胶条件的响应面优化 2.4.1 响应面优化试验结果以水料比、初始pH值和脱胶温度作为独立自变量,脱胶率为响应值,响应面试验结果见表 4。
| 实验编号 Experiment No. |
水料比 Water-to-material ratio |
初始pH值 Initial pH |
温度 Temperature (℃) |
脱胶率 Degumming rate (%) |
| 1 | 16:1 | 6.0 | 42 | 14.01 |
| 2 | 12:1 | 6.0 | 42 | 12.68 |
| 3 | 12:1 | 7.0 | 37 | 13.89 |
| 4 | 16:1 | 5.0 | 37 | 14.55 |
| 5 | 14:1 | 6.0 | 37 | 14.67 |
| 6 | 14:1 | 5.0 | 42 | 12.01 |
| 7 | 14:1 | 6.0 | 37 | 14.32 |
| 8 | 14:1 | 6.0 | 37 | 14.65 |
| 9 | 14:1 | 7.0 | 42 | 12.82 |
| 10 | 14:1 | 6.0 | 37 | 14.33 |
| 11 | 12:1 | 6.0 | 32 | 11.65 |
| 12 | 14:1 | 5.0 | 32 | 11.64 |
| 13 | 12:1 | 5.0 | 37 | 11.31 |
| 14 | 14:1 | 6.0 | 37 | 13.89 |
| 15 | 16:1 | 7.0 | 37 | 14.22 |
| 16 | 16:1 | 6.0 | 32 | 12.81 |
| 17 | 14:1 | 7.0 | 32 | 13.49 |
利用Design Expert对试验数据进行回归拟合分析,得到的回归方程模型如下:
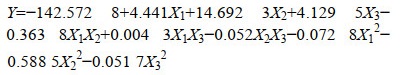
|
其中,X1-X3分别代表水料比、初始pH值和温度。方程式中各系数的正负可反映它们对苎麻脱胶率的影响方向,而系数绝对值的大小则可反映对苎麻脱胶率的影响程度。
对回归方程模型进行方差分析,结果见表 5。
| 来源 Source |
总和 Sum of squares |
自由度 Degree of freedom |
均方 Mean square |
F值 F value |
P值 P value |
| 模型 Model |
19.991 9 | 9 | 2.221 3 | 11.351 4 | 0.002 1** |
| X1:水料比 X1: Water-to-material ratio |
4.590 5 | 1 | 4.590 5 | 23.458 2 | 0.001 9** |
| X2:初始pH值 X2: Initial pH |
3.013 5 | 1 | 3.013 5 | 15.399 7 | 0.005 7** |
| X3:温度 X3: Temperature |
0.465 6 | 1 | 0.465 6 | 2.379 4 | 0.166 9 |
| X1X2 | 2.117 0 | 1 | 2.117 0 | 10.818 5 | 0.013 3* |
| X1X3 | 0.007 2 | 1 | 0.007 2 | 0.036 9 | 0.853 1 |
| X2X3 | 0.270 4 | 1 | 0.270 4 | 1.381 8 | 0.278 2 |
| X12 | 0.356 6 | 1 | 0.356 6 | 1.822 1 | 0.219 1 |
| X22 | 1.458 2 | 1 | 1.458 2 | 7.451 9 | 0.029 3* |
| X32 | 7.044 8 | 1 | 7.044 8 | 36.000 5 | 0.000 5** |
| 残差 Residual |
1.369 8 | 7 | 0.195 7 | ||
| 失拟项 Lack of fit |
0.966 9 | 3 | 0.322 3 | 3.200 0 | 0.145 3 |
| 净误差 Pure error |
0.402 9 | 4 | 0.100 7 | ||
| 总离差 Cor total |
21.361 7 | 16 | |||
| 注:R2=0.935 9;P > 0.1,差异不显著;*:P < 0.05,差异显著;**:P < 0.01,差异极显著 Note: R2=0.935 9;P > 0.1, Insignificant difference; *: P < 0.05, Significant difference; **: P < 0.01, Highly significant difference | |||||
由表 5中F值可知,各考察因素对苎麻脱胶率的影响因素次序是X1 (水料比) > X2 (初始pH) > X3 (温度)。该模型P值为0.002 1,小于0.01,差异极显著;失拟项P值为0.145 3,大于0.1,不显著;模型的决定系数R2=0.935 9,表明模型拟合度高,其可信性高。此外,一次项中X1、X2极显著,X3不显著;交互项中X1X2显著,X1X3和X2X3不显著;二次项中X12不显著,X22显著,X32极显著。
2.4.3 响应面分析为了分析回归方程预测的三因素对苎麻脱胶率的影响,利用Design-Expert得到响应曲面图(图 5-7)。由图 5-7可知,在一定范围内,脱胶率随水料比和初始pH值的增加而增加,高水料比低pH值和低水料比高pH值都不能获得较好的脱胶率,说明水料比和初始pH值对于菌株HG-9和HG-25混合脱胶有显著的交互作用。然而对于水料比与温度及初始pH值与温度,它们的交互作用对苎麻韧皮脱胶率的影响较小。

|
| 图 5 水料比和初始pH对脱胶率交互影响的响应面图(A)和等高线(B) Figure 5 Response surface plot (A) and contour line (B) of the interactive effect of water-to-material ratio and initial pH on the degumming rate |
|
|

|
| 图 6 水料比和温度对脱胶率交互影响的响应面图(A)和等高线(B) Figure 6 Response surface plot (A) and contour line (B) of the interactive effect of water-to-material ratio and temperature on degumming rate |
|
|

|
| 图 7 初始pH和温度对脱胶率交互影响的响应面图(A)和等高线(B) Figure 7 Response surface plot (A) and contour line (B) of the interactive effect of initial pH and temperature on the degumming rate |
|
|
利用Design Expert对菌株HG-9和HG-25混合脱胶条件进行优化,发现水料比16:1,初始pH值5.88,温度37.61 ℃时理论脱胶率最高,为14.86%。为了验证模型的可靠性,在上述条件下进行了苎麻脱胶试验。结合实际操作情况,水料比16:1保持不变,初始pH值调整为5.9,脱胶温度调整为37.6 ℃。结果表明脱胶率为14.82%±0.15%,同理论值仅相差0.04%,表明该试验模型的准确性。
在该优化条件下,苎麻的胶质去除率为52.91%,相对HG-9单独脱胶增加了9.32%,这一结果证明了2株菌具有很好的协同作用,即3种关键脱胶酶果胶酶、甘露聚糖酶和木聚糖酶对于苎麻胶质的去除十分关键,而且它们之间存在很好的协同作用,只用一种酶或者2种酶处理,均达不到很好的脱胶效果。刘唤明等[22]将果胶酶、甘露聚糖酶和木聚糖酶用于苎麻脱胶,分别用3种单一酶脱胶及3种酶联合脱胶,发现只用一种酶处理时,胶质去除率分别为4.3%、1.0%和1.2%;3种酶联合处理时最终残胶率为5.2%,进一步证实了3种酶间协同作用的存在。
2.5 脱胶时间对苎麻脱胶率的影响对于苎麻脱胶,尤其是大型脱胶系统,脱胶时间对其运行成本和脱胶效果有很大影响,基于菌株HG-9和HG-25的脱胶条件,对混合脱胶时间进行优化(图 8)。

|
| 图 8 脱胶时间对混合菌株HG-9/HG-25苎麻脱胶率的影响 Figure 8 Effect of degumming time on the degumming rate of ramie fibers by strain HG-9/HG-25 |
|
|
由图 8可以看出,随着脱胶时间的延长,苎麻脱胶率一直在增加,但14 h后增长缓慢。脱胶前期,菌株生长相对缓慢,分泌的酶无法满足对苎麻胶质的降解,而到达14 h后,微生物生长进入平稳期甚至衰亡期,残留的胶质与纤维素的结合更紧密,难以去除,导致脱胶效率降低。因此,最佳脱胶时间选择为14 h,比菌株HG-9单独脱胶缩短了2 h。
2.6 复合微生物脱胶效果评价 2.6.1 苎麻胶质去除率分析对苎麻原麻及不同微生物处理的苎麻进行总胶质、脂蜡质、水溶物、果胶、半纤维素和木质素的含量测定,结果如图 9所示。

|
| 图 9 苎麻组成成分含量分析 Figure 9 Analysis of ramie composition content |
|
|
从图 9可知,混合菌脱胶与HG-9单独脱胶相比,残留的脂蜡质和水溶物均有少量增加,而在纤维的纺纱过程中,一定量脂蜡质的存在是必需的;对于半纤维素,其含量由11.83%降至8.88%,去除率提高21.24%。菌株HG-9和HG-25并不分泌木质素降解酶类,但混合脱胶下木质素去除率增加17.93%,而木质素通过共价键与半纤维素连接,说明半纤维素可带动去除木质素。半纤维素和木质素去除率的增高可减少后期碱液和漂白剂的用量,具有显著的经济效益和环保效益。但是,HG-9/HG-25混合脱胶后,苎麻中主要的胶质成分,如半纤维素和果胶,并没有被完全去除。半纤维素和果胶是苎麻胶质的主要成分,分别占胶质总量的50%和17%[23]。与纤维素不同的是,果胶和半纤维素结构复杂,它们除了由1, 4-糖苷键形成的主链外,还带有很多支链,如葡萄糖醛酸、阿拉伯呋喃糖苷、乙酰基和半乳糖等[24-26]。这些支链结构的存在会造成空间障碍,降低主链降解酶和主链的可及性,限制降解酶的作用,导致果胶和半纤维素的不完全去除[27]。此外,在脱胶条件优化时,通气情况也会对菌株的生长和脱胶效果有一定的影响,在今后的脱胶研究中,一方面可引入这些支链降解酶与主链降解酶发挥协同作用,另一方面可通过对转速条件进行优化,从而进一步提高苎麻脱胶效率,减少后期化学试剂的使用,促进苎麻绿色高效脱胶。
2.6.2 漂白剂使用量分析以苎麻残胶率为指标,对菌株HG-9单独脱胶及其与HG-25混合脱胶后所需漂白剂用量进行对比(图 10)。

|
| 图 10 苎麻脱胶后期漂白剂使用量 Figure 10 Usage of bleacher in the later stage of ramie degumming |
|
|
由图 10可知,对于HG-9单独处理后的苎麻纤维,当次氯酸钠使用量为0.12 g时,其残胶率为1.70%,进一步增加次氯酸钠的用量,其残胶率几乎保持不变;而对于混合脱胶后的苎麻纤维,次氯酸钠使用量为0.10 g时,残胶率便可达到1.70%,增加次氯酸钠使用量,残胶率也不会进一步降低。对于国家一等品苎麻精干麻,其残胶率要低于2%,因此,对于混合脱胶后的苎麻纤维,只需0.08 g的次氯酸钠就可达到一等品要求,减少了次氯酸钠的使用量。这不仅可以减少对苎麻纤维的损伤也可降低水污染,具有很好的经济效益和环保效益。
2.6.3 苎麻纤维微观形态分析扫描电镜可用来观察苎麻纤维表面的形态结构。对苎麻原麻、菌株HG-9和HG-25单独脱胶及HG-9/HG-25混合脱胶后的样品进行扫描电镜测试(图 11)。

|
| 图 11 苎麻样品扫描电镜图 Figure 11 Scanning electron micrograph of ramie fibers 注:A:苎麻原麻,4 000×;B:菌株HG-9单独脱胶后的苎麻样品,2 400×;C:菌株HG-25单独脱胶后的苎麻样品,1 000×;D:混合菌脱胶后的苎麻样品,2 400× Note: A: The raw ramie fibers, 4 000×; B: The ramie fibers treated with HG-9, 2 400×; C: The ramie fibers treated with HG-25, 1 000×; D: The ramie fibers treated with HG-9/HG-25, 2 400× |
|
|
图 11A所示为苎麻原麻,可以看到,纤维之间存在很多胶质并粘连成纤维束,纤维束表面凹凸不平,存在网格状结构。图 11B和图 11C所示分别为单独使用菌株HG-9和HG-25脱胶后的苎麻纤维,可见纤维与之间的胶连物大大减少,单根纤维被暴露,但纤维表面还残留一些附着物。图 11D所示为混合菌脱胶后的苎麻纤维,与HG-9和HG-25单独脱胶相比,其纤维表面更加光滑平整,更多的胶质被去除,从微观角度证明了混合菌之间良好的协同作用。
3 结论本研究利用正交设计及响应面法对菌株HG-9和HG-25混合脱胶条件进行了优化,当接种量、水料比和初始pH值分别为6%、16:1和5.9时,在37.6 ℃下处理14 h,脱胶效果最佳,而且二者在脱胶过程中具有很好的协同作用。与菌株HG-9单独脱胶相比,脱胶时间缩短2 h,胶质去除率提高9.32%,漂白剂用量减少20%,而且纤维表面变得更加光滑、平整,纤维分散度更好。由此可见,果胶酶、甘露聚糖酶和木聚糖酶具有很好的协同作用,并在苎麻脱胶中发挥重要的作用。本研究不仅提高苎麻胶质去除率,缩短苎麻脱胶时间,而且减少脱胶后期漂白剂用量,为苎麻绿色生物脱胶的进一步优化及工业化应用提供了指导。
| [1] |
Ni JL, Zhu AG, Wang XF, Xu Y, Sun ZM, Chen JH, Luan MB. Genetic diversity and population structure of ramie (Boehmeria nivea L.)[J]. Industrial Crops and Products, 2018, 115: 340-347. DOI:10.1016/j.indcrop.2018.01.038 |
| [2] |
Sarkar D, Sinha MK, Kundu A, Kar C. Why is ramie the strongest yet stiffest of bast fibers[J]. Current Science, 2010, 98(12): 1570-1572. |
| [3] |
Mukhopadhyay A, Dutta N, Chattopadhyay D, Chakrabarti K. Degumming of ramie fiber and the production of reducing sugars from waste peels using nanoparticle supplemented pectate lyase[J]. Bioresource Technology, 2013, 137: 202-208. DOI:10.1016/j.biortech.2013.03.139 |
| [4] |
Mao KW, Chen HG, Qi HH, Qiu ZD, Zhang L, Zhou JG. Visual degumming process of ramie fiber using a microbial consortium RAMCD407[J]. Cellulose, 2019, 26(5): 3513-3528. DOI:10.1007/s10570-019-02288-1 |
| [5] |
Cheng LF, Liu ZC, Feng XY, Wang QM, Li Q, Zheng K, Yang Q, Duan SW. Screening on compound pectinase for ramie degumming and its effect analysis[J]. Journal of Textile Research, 2017, 38(6): 64-68. (in Chinese) 成莉凤, 刘正初, 冯湘沅, 汪启明, 李琦, 郑科, 杨琦, 段盛文. 苎麻脱胶果胶复合酶的优选及其效果分析[J]. 纺织学报, 2017, 38(6): 64-68. |
| [6] |
Fan XS, Liu ZW, Liu ZT, Lu J. A novel chemical degumming process for ramie bast fiber[J]. Textile Research Journal, 2010, 80(19): 2046-2051. DOI:10.1177/0040517510373632 |
| [7] |
Guo FF, Zou MY, Li XZ, Zhao J, Qu YB. An effective degumming enzyme from Bacillus sp. Y1 and synergistic action of hydrogen peroxide and protease on enzymatic degumming of ramie fibers[J]. BioMed Research International, 2013, 2013: 212315. |
| [8] |
Liu ZC, Duan SW, Sun QX, Peng YD, Feng XY, Zheng K, Hu ZX, Zhang YX. A rapid process of ramie bio-degumming by Pectobacterium sp. CXJZU-120[J]. Textile Research Journal, 2012, 82(15): 1553-1559. DOI:10.1177/0040517512444338 |
| [9] |
Lyu ZR, Li ZF, Jiang HL, Sun ZY. Research progress of ramie fiber preparation[J]. Cotton Textile Technology, 2020, 48(11): 68-72. (in Chinese) 吕泽瑞, 黎征帆, 江惠林, 孙泽宇. 苎麻纤维制备研究进展[J]. 棉纺织技术, 2020, 48(11): 68-72. DOI:10.3969/j.issn.1001-7415.2020.11.020 |
| [10] |
Wang Q, Chen HG, Liu L, Guo SW, Liu JS. Isolation, identification and degumming efficiency of dominant bacteria from ramie degumming bacterial consortium RAMCD407[J]. Microbiology China, 2016, 43(12): 2561-2567. (in Chinese) 王茜, 陈洪高, 刘露, 郭帅威, 柳建设. 苎麻脱胶菌群RAMCD407中优势菌的分离、鉴定及脱胶能力分析[J]. 微生物学通报, 2016, 43(12): 2561-2567. |
| [11] |
Fan P, He F, Yang Y, Ao MZ, Ouyang J, Liu Y, Yu LJ. In-situ microbial degumming technology with Bacillus sp. HG-28 for industrial production of ramie fibers[J]. Biochemical Engineering Journal, 2015, 97: 50-58. DOI:10.1016/j.bej.2014.12.010 |
| [12] |
Cheng LF, Wang QM, Feng XY, Duan SW, Yang Q, Zheng K, Liu ZY, Liu ZC, Peng YD. Screening a bacterium and its effect on the biological degumming of ramie and kenaf[J]. Scientia Agricola, 2018, 75(5): 375-380. DOI:10.1590/1678-992x-2017-0108 |
| [13] |
Shu T, Bai Y, Wang YW, Wang HH, Li PD, Xiang MX, Yu TY, Xu H, Yu LJ. A high-efficiency and eco-friendly degumming process for ramie fibers[J]. Journal of Cleaner Production, 2020, 276: 124217. DOI:10.1016/j.jclepro.2020.124217 |
| [14] |
Basu S, Saha MN, Chattopadhyay D, Chakrabarti K. Large-scale degumming of ramie fibre using a newly isolated Bacillus pumilus DKS1 with high pectate lyase activity[J]. Journal of Industrial Microbiology & Biotechnology, 2009, 36(2): 239-245. |
| [15] |
Duan SW, Liu ZC, Feng XY, Zheng K, Cheng LF, Hu ZX. Study on the diversity of the herbaceous fiber extraction strain resources[J]. Plant Fiber Sciences in China, 2007, 29(6): 330-333. (in Chinese) 段盛文, 刘正初, 冯湘沅, 郑科, 成莉凤, 胡镇修. 草本纤维提取菌种资源多样性研究[J]. 中国麻业科学, 2007, 29(6): 330-333. DOI:10.3969/j.issn.1671-3532.2007.06.007 |
| [16] |
Xiao L, Wang GX, Chen GJ. The research advancement of the enzymatic degumming of ramie (Boehmeria nivea)[J]. Microbiology China, 2004, 31(5): 101-105. (in Chinese) 肖丽, 王贵学, 陈国娟. 苎麻酶法脱胶的研究进展[J]. 微生物学通报, 2004, 31(5): 101-105. DOI:10.3969/j.issn.0253-2654.2004.05.022 |
| [17] |
Zheng LS, Du YM, Zhang JY. Degumming of ramie fibers by alkalophilic bacteria and their polysaccharide-degrading enzymes[J]. Bioresource Technology, 2001, 78(1): 89-94. DOI:10.1016/S0960-8524(00)00154-1 |
| [18] |
Du ZF, Huang FR. Process optimization of composite microorganism degumming of ramie[J]. Journal of Textile Research, 2012, 33(5): 56-61. (in Chinese) 杜兆芳, 黄芙蓉. 苎麻复合微生物脱胶工艺优化[J]. 纺织学报, 2012, 33(5): 56-61. DOI:10.3969/j.issn.0253-9721.2012.05.012 |
| [19] |
Wang Q, Chen HG, Fang G, Chen AQ, Yuan P, Liu JS. Isolation of Bacillus cereus P05 and Pseudomonas sp. X12 and their application in the ramie retting[J]. Industrial Crops and Products, 2017, 97: 518-524. DOI:10.1016/j.indcrop.2016.12.047 |
| [20] |
Wang TL, Qiu HW, Chen HH, An Y, Ding LX. Applying 3, 5-dinitrosalicylic acid method to measure pectinase activity[J]. Food & Machinery, 2008, 24(3): 96-99, 104. (in Chinese) 王天龙, 仇宏伟, 陈海华, 安燕, 丁立孝. 3, 5-二硝基水杨酸法测定果胶酶活力的条件研究[J]. 食品与机械, 2008, 24(3): 96-99, 104. |
| [21] |
National Standard (Mandatory) of the People's Republic of China: method of quantitative analysis of ramie chemical components. GB 5889—1986[S]. Beijing: Standards Press of China, 1986(in Chinese) 国家标准局. 中华人民共和国国家标准: 苎麻化学成分定量分析方法GB 5889—1986[S]. 北京: 中国标准出版社, 1986 |
| [22] |
Liu HM, Liang YX, Peng DX. Study on the the enzymatic degumming of ramie[J]. Plant Fibers and Products, 2006, 28(2): 87-90. (in Chinese) 刘唤明, 梁运祥, 彭定祥. 苎麻酶法脱胶的研究[J]. 中国麻业, 2006, 28(2): 87-90. DOI:10.3969/j.issn.1671-3532.2006.02.008 |
| [23] |
Li ZF, Li ZL, Ding RY, Yu CW. Composition of ramie hemicelluloses and effect of polysaccharides on fiber properties[J]. Textile Research Journal, 2016, 86(5): 451-460. DOI:10.1177/0040517515592811 |
| [24] |
Round AN, Rigby NM, MacDougall AJ, Morris VJ. A new view of pectin structure revealed by acid hydrolysis and atomic force microscopy[J]. Carbohydrate Research, 2010, 345(4): 487-497. DOI:10.1016/j.carres.2009.12.019 |
| [25] |
She YL, Li XT, Song HL, Ma JJ, Lyu YG. Research advance in microorganism xylanases[J]. China Brewing, 2009, 28(2): 1-4. (in Chinese) 佘元莉, 李秀婷, 宋焕禄, 马家津, 吕跃钢. 微生物木聚糖酶的研究进展[J]. 中国酿造, 2009, 28(2): 1-4. DOI:10.3969/j.issn.0254-5071.2009.02.001 |
| [26] |
Moreira LRS, Filho EXF. An overview of mannan structure and mannan-degrading enzyme systems[J]. Applied Microbiology and Biotechnology, 2008, 79(2): 165-178. DOI:10.1007/s00253-008-1423-4 |
| [27] |
Zhu TD, He Y, Yang B, Liu JC, Wu MC. Combinative degradation of xylan with acetyl xylan esterase and xylanase[J]. Journal of Food Science and Biotechnology, 2016, 35(4): 375-380. (in Chinese) 朱天地, 何瑶, 杨标, 刘加驰, 邬敏辰. 乙酰木聚糖酯酶协同木聚糖酶降解木聚糖的研究[J]. 食品与生物技术学报, 2016, 35(4): 375-380. DOI:10.3969/j.issn.1673-1689.2016.04.007 |