 2019, Vol. 46
2019, Vol. 46扩展功能
文章信息
- 吴楠, 田风华, 贾传文, 王悦, 李长田
- WU Nan, TIAN Feng-Hua, JIA Chuan-Wen, WANG Yue, LI Chang-Tian
- 混料设计优化红平菇菌丝生长的“以草代木”配方
- Substrate optimization for mycelium growth of Pleurotus djamor in the concept of "replacing wood by grass"
- 微生物学通报, 2019, 46(6): 1390-1403
- Microbiology China, 2019, 46(6): 1390-1403
- DOI: 10.13344/j.microbiol.china.180509
-
文章历史
- 收稿日期: 2018-07-02
- 接受日期: 2018-09-10
- 网络首发日期: 2018-09-29






2. 吉林工程职业学院 吉林 长春 130000
2. Jilin Engineering Vocational College, Changchun, Jilin 130000, China
现如今人们越来越重视生态环境的保护,树木的采伐受到控制,自1998年天然林保护工程实施以来,木屑等原材料日趋紧张,导致生产成本逐年增加[1]。传统食药用菌生产主要以林木为原料,菌业的快速发展导致“菌林矛盾”日益突出。因此,“以草代木”已经成为当今食用菌产业研究的热点,用这种方式生产食药用菌与传统栽培的食药用菌相比,具有活性成分含量高、易于分离的特点[2]。红平菇Pleurotus djamor (Rumph.) Boedijn是一种生长在热带、亚热带地区的木腐菌类,子实体色泽鲜艳、味道鲜美、营养丰富,属低脂肪高蛋白食品,含多种人体所需氨基酸、维生素及矿物质,是一种“能吃能观赏”的珍稀蕈菌[3-6],其提取物对癌细胞有抑制作用,锌多糖可有效减轻肝脏和肾脏的损伤[7-8]。
红平菇生长过程中,各种酶在其菌丝体发育中起到不可或缺的作用。门金鑫等[9]以生物量、纤维素酶酶活、漆酶酶活等为指标筛选出纤维素酶、漆酶活性高的优质蜜环菌菌株用于猪苓栽培。张政研究发现,当草菇菌丝体退化时间延长,多酚氧化酶、漆酶等酶活呈现明显降低趋势[10]。漆酶和多酚氧化酶都是木质素降解的重要催化剂,可以将木质素芳香族高分子化合物分解为小分子物质供菌丝生长,胞外漆酶活性越强,菌株降解木质素的能力就越强[10-13]。纤维素酶是一种复合酶,是降解基质纤维素的主要酶类,由羧甲基纤维素酶和滤纸纤维素酶等酶组成,可将纤维素分解成小分子物质供菌丝体生长发育使用[14]。
关于农业剩余物基质代谢的研究,国内有许多报道,如胡居吾[15]、冯作山[16]、刘连强[17]、张鹏等[18]先后对灵芝、白灵菇、金针菇进行了“以草代木”栽培基质的研究。目前,红平菇降解农业剩余物代谢基质的研究鲜有报道,因此有必要对适合红平菇菌丝生长的“农业剩余物”基质组分进行研究。
本试验以7种农业剩余物做主料,利用软件Design-Expert 8.0.6.1设计基质配方培养红平菇,探究不同基质组分下其胞外酶活性及菌丝生长速率的差异,以期筛选出适合红平菇菌丝生长的“以草代木”基质配方。
1 材料与方法 1.1 材料 1.1.1 菌株和样品 红平菇,编号2014062001,由吉林农业大学食药用菌种保藏中心提供。小麦秸秆、水稻秸秆、大豆秸秆、玉米秸秆、玉米芯、花生秸秆、油菜秸秆、杨木屑、白糖、麦麸、石膏粉,由吉林农业大学菌菜基地提供。
1.1.2 主要试剂和仪器 柠檬酸、柠檬酸钠、3, 5-二硝基水杨酸(DNS)、联苯胺、醋酸、醋酸钠、磷酸二氢钠、磷酸氢二钠、邻苯二酚,长春鑫海泰生物科技有限公司。生化培养箱,宁波赛福实验仪器有限公司;多功能酶标仪,BioTek公司。 1.2 方法 1.2.1 初筛培养基的制备 分别称取7种秸秆粉各50 g,代替基础配方(木屑78.0%,麦麸20.0%,石灰粉1.0%,蔗糖1.0%)中的木屑,其他辅料成分不变,等量分装于10个平板(直径80 mm)中,含水量78%,1×105 Pa灭菌40 min,冷却至室温备用。 1.2.2 菌丝生长速率测定 使用5 mm口径的打孔器,沿长满菌丝的PDA平板边缘均匀打孔,将菌块接种至培养基中心,24 ℃黑暗条件下培养6 d,每种培养基5个重复。每隔24 h采用十字画线法测得当天菌丝生长速率,最后计算平均生长速率,即菌丝生长速率。 1.2.3 粗酶液的提取[19] 准确称取在平板中24 ℃黑暗条件下培养至第6天的菌质2 g,适当研磨后移入试管中,加纯净水20 mL,于37 ℃水浴锅中浸提2 h后,4层纱布过滤,取滤液,定容至25 mL,4 ℃、8 000 r/min冷冻离心10 min,上清液即为粗酶液,5 mL离心管分装,于−80 ℃保存备用。 1.2.4 酶活的测定 漆酶、多酚氧化酶酶活的测定参照韩增华等的方法。漆酶酶活力单位定义:以每毫升培养酶液与底物作用30 min内改变0.1光密度值为一个活力单位(U)。多酚氧化酶酶活力单位定义:以每毫升培养酶液与底物作用30 min内改变0.1光密度值为一个活力单位(U)[14]。纤维素酶酶活的测定参照安琪等[20]的方法。纤维素酶酶活力单位定义为:每分钟催化底物生成1 μmol葡萄糖所需的酶量为一个酶活单位(U)[20]。 1.2.5 混合设计配方 采用混料设计(Mixture)中的限制成分上下界的单纯型格子法(Simplex-lattice design)进行设计,利用软件Design-Expert 8.0.6.1对主料中各成分的比例进行优化设计,X1–X5均设在0–100%,X为主料,主料成分占整个培养基的75%,其余辅料麦麸、玉米面、豆粕、石灰粉、石膏粉占25%。X1为小麦秸秆、X2为油菜秸秆、X3为花生秸秆、X4为大豆秸秆、X5为水稻秸秆。X1+X2+X3+X4+X5=100%。对照配方:玉米芯45.0%,木屑30.0%,麦麸15.0%,玉米面5.0%,豆粕3.0%,石灰1.0%,石膏1.0% (表 1)。| 配方 Formulation |
基质配比Substrate ratio (%) | ||||
| X1 | X2 | X3 | X4 | X5 | |
| 1 | 100 | 0 | 0 | 0 | 0 |
| 2 | 0 | 100 | 0 | 0 | 0 |
| 3 | 0 | 0 | 100 | 0 | 0 |
| 4 | 0 | 0 | 0 | 100 | 0 |
| 5 | 0 | 0 | 0 | 0 | 100 |
| 6 | 50 | 50 | 0 | 0 | 0 |
| 7 | 50 | 0 | 50 | 0 | 0 |
| 8 | 50 | 0 | 0 | 50 | 0 |
| 9 | 50 | 0 | 0 | 0 | 50 |
| 10 | 0 | 50 | 50 | 0 | 0 |
| 11 | 0 | 50 | 0 | 50 | 0 |
| 12 | 0 | 50 | 0 | 0 | 50 |
| 13 | 0 | 0 | 50 | 50 | 0 |
| 14 | 0 | 0 | 50 | 0 | 50 |
| 15 | 0 | 0 | 0 | 50 | 50 |
| 16 | 60 | 10 | 10 | 10 | 10 |
| 17 | 10 | 60 | 10 | 10 | 10 |
| 18 | 10 | 10 | 60 | 10 | 10 |
| 19 | 10 | 10 | 10 | 60 | 10 |
| 20 | 10 | 10 | 10 | 10 | 60 |
| 21 | 20 | 20 | 20 | 20 | 20 |
| 注:X1:小麦秸秆、X2:油菜秸秆、X3:花生秸秆、X4:大豆秸秆、X5:水稻秸秆. Note: X1: Wheat straw; X2: Rape straw; X3: Peanut straw; X4: Soybean straw; X5: Rice straw. |
|||||
根据7种农业剩余物及木屑的平板培养试验,测得菌丝生长速率做差异显著性分析,发现小麦秸秆做基质菌丝生长速率最高,为6.76 mm/d,其次是大豆秸秆和花生秸秆,分别为6.10 mm/d和5.70 mm/d。木屑做主料时菌生长速率是5.59 mm/d,与油菜秸秆(5.62 mm/d)、水稻秸秆(5.62 mm/d)和玉米秸秆(5.51 mm/d)无差异显著性,但在数值上玉米秸秆较低。玉米芯做基质时菌丝生长速率最低,为5.40 mm/d。因此筛选出小麦秸秆、大豆秸秆、花生秸秆、油菜秸秆和水稻秸秆5种优势主料(图 1)。

|
| 图 1 初筛配方红平菇菌丝生长速率差异显著性分析 Figure 1 Analysis on the difference of growth rate of mycelium of Pleurotus djamor with initial screening formula 注:图中不同字母代表差异性显著,相同字母代表差异性不显著. Note: The different letters represent significant difference, and the same letters represent no significant difference. |
|
|

|
通过分析菌丝生长速率的二次多项回归模型方差(表 2),结果显示:线性混合模型和二次回归模型的P值都小于0.000 1,二者都达到了极显著水平,相关系数R2=0.894 3,很好地拟合了主料与菌丝生长速率的关系。失拟项小于0.000 1是极显著的,表明失拟误差除了与试验误差有关,还与其他因素呈极显著相关。在回归模型中,方程系数K(X4)=6.39 > K(X1)=6.06 > K(X5)=5.83 > K(X3)=5.72 > K(X2)=4.95,表明各成分对菌丝生长速率的贡献程度为:X4 (大豆秸秆) > X1 (小麦秸秆) > X5 (水稻秸秆) > X3 (花生秸秆) > X2 (油菜秸秆)。交互作用X1X2、X1X3和X2X4是极显著的,表明小麦秸秆与油菜秸秆、小麦秸秆与花生秸秆、油菜秸秆与大豆秸秆交互作用可以显著影响红平菇菌丝的生长。做22个配方7 d菌丝生长直径的差异显著性分析,结果表明:4、7、9、15、18、19号配方与对照配方的菌丝生长速率无显著差异,20、21号配方菌丝生长速率与对照配方有差异显著性并高于对照配方;10、11号配方菌丝生长最慢(图 2)。如图 3所示,小麦秸秆与油菜秸秆、小麦秸秆与花生秸秆可显著提高菌丝生长速率。油菜秸秆与大豆秸秆交互作用不利于红平菇菌丝体的生长。一个配方的C/N会显著影响菌丝生长速率,如冯改静等研究发现适合猴头菌菌丝体生长的C/N范围为28:1-38:1[21]。根据表 3可以得知,不同的秸秆粉碳氮含量会有差异,因此每个配方的C/N也不相同,从而影响菌丝生长速率。
| 变异源 Source |
平方和 Sum of squares |
自由度 df |
均方 Mean square |
F | P |
| 模型 Model | 25.23 | 14 | 1.80 | 8.41 | < 0.000 1 |
| 线性混合模型 Linear mixed model | 14.50 | 4 | 3.62 | 16.92 | < 0.000 1 |
| X1X2 | 1.93 | 1 | 1.93 | 9.01 | 0.004 2 |
| X1X3 | 1.66 | 1 | 1.66 | 7.76 | 0.007 6 |
| X1X4 | 0.058 | 1 | 0.058 | 0.27 | 0.606 6 |
| X1X5 | 1.18 | 1 | 1.18 | 5.49 | 0.023 3 |
| X2X3 | 1.06 | 1 | 1.06 | 4.95 | 0.030 8 |
| X2X4 | 2.69 | 1 | 2.69 | 12.54 | 0.000 9 |
| X2X5 | 0.94 | 1 | 0.94 | 4.39 | 0.041 5 |
| X3X4 | 0.36 | 1 | 0.36 | 1.67 | 0.202 9 |
| X3X5 | 0.68 | 1 | 0.68 | 3.18 | 0.080 9 |
| X4X5 | 0.92 | 1 | 0.92 | 4.31 | 0.043 2 |
| 残差 Residual | 10.28 | 48 | 0.21 | ||
| 失拟项 Lack of fit | 7.90 | 6 | 1.32 | 23.20 | < 0.000 1 |
| 纯误差 Pure error | 2.38 | 42 | 0.057 | ||
| 总和 Cor total | 35.51 | 62 |

|
| 图 2 22个配方菌丝生长速率差异显著性分析 Figure 2 Analysis of the significant difference in the activity of mycelium growth rate in 22 formulations 注:图中不同字母代表差异性显著,相同字母代表差异性不显著. Note: The different letters represent significant difference, and the same letters represent no significant difference. |
|
|

|
| 图 3 各主料交互作用影响菌丝生长速率等高线图 Figure 3 The main interaction effect of the mycelium growth rate contour map 注:A:小麦秸秆、花生秸秆和油菜秸秆三者交互作用;B:小麦秸秆、大豆秸秆和油菜秸秆三者交互作用;C:小麦秸秆、水稻秸秆和油菜秸秆三者交互作用;D:油菜秸秆、花生秸秆和大豆秸秆三者交互作用;E:油菜秸秆、水稻秸秆、花生秸秆三者交互作用;F:花生秸秆、大豆秸秆和水稻秸秆三者交互作用;G:大豆秸秆、水稻秸秆和小麦秸秆三者交互作用. Note: A: The interaction of wheat straw, peanut straw and rape straw; B: The interaction of wheat straw, soybean straw and rape straw; C: The interaction of wheat straw, rice straw and rape straw; D: The interaction of rape straw, peanut straw and soybean straw; E: The interaction of rape straw, rice straw, peanut straw; F: The interaction of peanut straw, soybean straw and rice straw; G: The interaction of soybean straw, rice straw and wheat straw. |
|
|
| 成分 Component | 纤维素 Cellulose (%) | 半纤维素 Hemicellulose (%) | 木质素 Lignin (%) | C (%) | N (%) |
| 小麦秸秆 Wheat straw | 39.26 | 21.19 | 22.10 | 41.84 | 0.59 |
| 油菜秸秆 Rape straw | 42.02 | 15.27 | 20.08 | 42.75 | 0.69 |
| 花生秸秆 Peanut straw | 31.05 | 11.54 | 26.03 | 45.52 | 0.84 |
| 大豆秸秆 Soybean straw | 30.83 | 14.64 | 32.80 | 44.27 | 0.59 |
| 水稻秸秆 Rice straw | 41.08 | 18.54 | 19.31 | 41.02 | 0.59 |
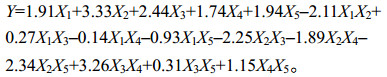
|
从其二次多项回归模型方差分析中可看出,二次多项回归模型和线性混合模型的方差均达到极显著水平,相关系数R2=0.923 0,很好地拟合了主料基质与多酚氧化酶活性之间的关系(表 4)。在回归方程中,方程系数K(X2)=3.33 > K(X3)=2.44 > K(X5)= 1.94 > K(X1)=1.91 > K (X4)=1.74,表明各成分对多酚氧化酶酶活的贡献程度为:X2 (油菜秸秆) > X3 (花生秸秆) > X5 (水稻秸秆) > X1 (小麦秸秆) > X4 (大豆秸秆)。根据表 4可知,交互作用X1X2、X2X3、X2X4、X2X5与X3X4是极显著的,表明其相互配比可以显著影响多酚氧化酶的活性。对这21个配方做差异显著性分析的结果(图 4)表明:2号配方的多酚氧化酶活性最高,4号配方多酚氧化酶活性最低。结合图 5可发现,小麦秸秆与油菜秸秆、油菜秸秆与花生秸秆、油菜秸秆与大豆秸秆、油菜秸秆与水稻秸秆交互作用可显著降低多酚氧化酶活性;花生秸秆与大豆秸秆交互作用可显著提高多酚氧化酶活性。从表 3可以看到,虽然油菜秸秆的多酚氧化酶活性最高,但是其木质素含量相对较少,由此可知除木质素外,还有其他底物能影响多酚氧化酶活性。
| 变异源 Source |
平方和 Sum of squares |
自由度 df |
均方 Mean square |
F | P |
| 模型 Model | 8.73 | 14 | 0.62 | 24.31 | < 0.000 1 |
| 线性混合模型 Linear mixed model | 5.02 | 4 | 1.25 | 48.91 | < 0.000 1 |
| X1X2 | 0.60 | 1 | 0.60 | 23.53 | < 0.000 1 |
| X1X3 | 9.990E-003 | 1 | 9.990E-003 | 0.39 | 0.535 5 |
| X1X4 | 2.583E-003 | 1 | 2.583E-003 | 0.10 | 0.752 3 |
| X1X5 | 0.12 | 1 | 0.12 | 4.59 | 0.037 2 |
| X2X3 | 0.68 | 1 | 0.68 | 26.57 | < 0.000 1 |
| X2X4 | 0.48 | 1 | 0.48 | 18.84 | < 0.000 1 |
| X2X5 | 0.74 | 1 | 0.74 | 28.81 | < 0.000 1 |
| X3X4 | 1.43 | 1 | 1.43 | 55.90 | < 0.000 1 |
| X3X5 | 0.013 | 1 | 0.013 | 0.50 | 0.484 7 |
| X4X5 | 0.18 | 1 | 0.18 | 6.94 | 0.011 3 |
| 残差 Residual | 1.23 | 48 | 0.026 | ||
| 失拟项 Lack of fit | 1.02 | 6 | 0.17 | 33.58 | < 0.000 1 |
| 纯误差 Pure error | 0.21 | 42 | 5.055E-003 | ||
| 总和 Cor total | 9.96 | 62 |

|
| 图 4 22个配方多酚氧化酶酶活差异显著性分析 Figure 4 Analysis of the significant difference in the activity of polyphenol oxidase in 22 formulations 注:图中不同小写字母代表差异性显著,相同字母代表差异性不显著 Note: The different letters represent significant difference, and the same letters represent no significant difference. |
|
|

|
| 图 5 各主料交互作用影响多酚氧化酶酶活等高线图 Figure 5 The main interaction effect of the activity of polyphenol oxidase contour map 注:A:小麦秸秆、花生秸秆和油菜秸秆三者交互作用;B:小麦秸秆、大豆秸秆和油菜秸秆三者交互作用;C:小麦秸秆、水稻秸秆和油菜秸秆三者交互作用;D:油菜秸秆、花生秸秆和大豆秸秆三者交互作用;E:油菜秸秆、水稻秸秆、花生秸秆三者交互作用;F:花生秸秆、大豆秸秆和水稻秸秆三者交互作用;G:大豆秸秆、水稻秸秆和小麦秸秆三者交互作用. Note: A: The interaction of wheat straw, peanut straw and rape straw; B: The interaction of wheat straw, soybean straw and rape straw; C: The interaction of wheat straw, rice straw and rape straw; D: The interaction of rape straw, peanut straw and soybean straw; E: The interaction of rape straw, rice straw, peanut straw; F: The interaction of peanut straw, soybean straw and rice straw; G: The interaction of soybean straw, rice straw and wheat straw. |
|
|
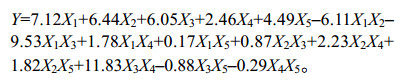
|
试验结果表明:二次多项回归模型和线性混合模型的方差均达到极显著水平,相关系数R2=0.982 5,很好地拟合了主料基质与纤维素酶活性之间的关系(表 5)。在回归模型中,方程系数K(X1)=7.12 > K(X2)=6.44 > K(X3)=6.05 > K(X5)=4.49 > K(X4)=2.46,表明各成分对纤维素酶酶活的贡献程度如下:X1 (小麦秸秆) > X2 (油菜秸秆) > X3 (花生秸秆) > X5 (水稻秸秆) > X4 (大豆秸秆)。交互作用X1X3、X3X4呈极显著水平,表明其在栽培料中相互配比可以显著影响纤维素酶活性(表 5)。做22个配方差异显著性分析可知,利用农业剩余物做主料的21个配方中,有17个配方的纤维素酶活性高于对照配方(图 6)。结合图 7可发现,小麦秸秆与花生秸秆交互作用可显著降低纤维素酶活性,花生秸秆与大豆秸秆交互作用可显著提高纤维素酶活性。由图 6和表 3可以发现,小麦秸秆、油菜秸秆纤维素含量较高,其纤维素酶活性也最高。而花生秸秆的纤维素和半纤维素含量最低,其纤维素酶酶活却高于和纤维素含量较高的水稻秸秆,由此说明花生秸秆中还有大量其他的纤维素酶降解的底物。
| 变异源 Source |
平方和 Sum of squares |
自由度 df |
均方 Mean square |
F | P |
| 模型 Model | 76.83 | 14 | 5.49 | 6.93 | < 0.000 1 |
| 线性混合模型 Linear mixed model | 35.53 | 4 | 8.88 | 11.22 | < 0.000 1 |
| X1X2 | 5.03 | 1 | 5.03 | 6.36 | 0.015 1 |
| X1X3 | 12.25 | 1 | 12.25 | 15.47 | 0.000 3 |
| X1X4 | 0.43 | 1 | 0.43 | 0.54 | 0.465 1 |
| X1X5 | 4.044E-003 | 1 | 4.044E-003 | 5.108E-003 | 0.943 3 |
| X2X3 | 0.10 | 1 | 0.10 | 0.13 | 0.720 4 |
| X2X4 | 0.67 | 1 | 0.67 | 0.85 | 0.361 9 |
| X2X5 | 0.45 | 1 | 0.45 | 0.56 | 0.456 6 |
| X3X4 | 18.90 | 1 | 18.90 | 23.87 | < 0.000 1 |
| X3X5 | 0.10 | 1 | 0.10 | 0.13 | 0.717 5 |
| X4X5 | 0.011 | 1 | 0.011 | 0.014 | 0.905 8 |
| 残差 Residual | 38.00 | 48 | 0.79 | ||
| 失拟项 Lack of fit | 37.25 | 6 | 6.21 | 350.71 | < 0.000 1 |
| 纯误差 Pure error | 0.74 | 42 | 0.018 | ||
| 总和 Cor total | 114.83 | 62 |

|
| 图 6 22个配方纤维素酶酶活差异显著性分析 Figure 6 Analysis of the significant difference in the activity of cellulase in 22 formulations 注:图中不同小写字母代表差异性显著,相同字母代表差异性不显著. Note: The different letters represent significant difference, and the same letters represent no significant difference. |
|
|

|
| 图 7 各主料交互作用影响纤维素酶酶活等高线图 Figure 7 The main interaction effect of the activity of cellulase contour map 注:A:小麦秸秆、花生秸秆和油菜秸秆三者交互作用;B:小麦秸秆、大豆秸秆和油菜秸秆三者交互作用;C:小麦秸秆、水稻秸秆和油菜秸秆三者交互作用;D:油菜秸秆、花生秸秆和大豆秸秆三者交互作用;E:油菜秸秆、水稻秸秆、花生秸秆三者交互作用;F:花生秸秆、大豆秸秆和水稻秸秆三者交互作用;G:大豆秸秆、水稻秸秆和小麦秸秆三者交互作用. Note: A: The interaction of wheat straw, peanut straw and rape straw; B: The interaction of wheat straw, soybean straw and rape straw; C: The interaction of wheat straw, rice straw and rape straw; D: The interaction of rape straw, peanut straw and soybean straw; E: The interaction of rape straw, rice straw, peanut straw; F: The interaction of peanut straw, soybean straw and rice straw; G: The interaction of soybean straw, rice straw and wheat straw. |
|
|
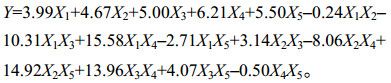
|
试验结果表明:二次多项回归模型和线性混合模型的方差均呈极显著水平,相关系数R2=0.995 8,表明方程模型与试验数据有99.58%的符合度,模型可信度较高,很好地拟合了各主料基质与漆酶活性之间的关系(表 6)。在回归模型中,方程系数K(X4)=6.21 > K(X5)= 5.50 > K(X3)= 5.00 > K(X2)= 4.67 > K(X1)=3.99,表明各成分对漆酶酶活的贡献程度为:X4 (大豆秸秆) > X5 (水稻秸秆) > X3 (花生秸秆) > X2 (油菜秸秆) > X1 (小麦秸秆)。交互作用X1X3、X1X4、X2X5、X3X4是极显著的(表 6),表明其在栽培料中相互配比可以显著影响漆酶活性。做22个配方差异显著性分析可知,利用农业剩余物做主料的21个配方中,有5个配方的漆酶活性高于对照配方(图 8)。结合图 9可看出,小麦秸秆与花生秸秆交互作用可显著降低胞外漆酶活性;小麦秸秆与大豆秸秆、油菜秸秆与水稻秸秆、花生秸秆与大豆秸秆交互作用可显著提高胞外漆酶活性。由图 8和表 3可以发现,漆酶活性在5种秸秆粉单独做主料时,与木质素含量呈显著的正相关。但当各主料混合配比时,漆酶酶活与木质素含量之间无相关性。由此可知,木质素含量在单一主料中对漆酶活性影响较大,在复合主料中,木质素含量对漆酶活性影响较弱,推测不同的碳氮比会显著影响漆酶的活性。
| 变异源 Source |
平方和 Sum of squares |
自由度 df |
均方 Mean square |
F | P |
| 模型 Model | 193.36 | 14 | 13.81 | 9.42 | < 0.000 1 |
| 线性混合模型 Linear mixed model | 44.50 | 4 | 11.12 | 7.59 | < 0.000 1 |
| X1X2 | 7.861E-003 | 1 | 7.861E-003 | 5.363E-003 | 0.941 9 |
| X1X3 | 14.36 | 1 | 14.36 | 9.79 | 0.003 0 |
| X1X4 | 32.78 | 1 | 32.78 | 22.36 | < 0.000 1 |
| X1X5 | 0.99 | 1 | 0.99 | 0.67 | 0.415 6 |
| X2X3 | 1.33 | 1 | 1.33 | 0.91 | 0.345 7 |
| X2X4 | 8.76 | 1 | 8.76 | 5.98 | 0.018 2 |
| X2X5 | 30.06 | 1 | 30.06 | 20.50 | < 0.000 1 |
| X3X4 | 26.30 | 1 | 26.30 | 17.94 | 0.000 1 |
| X3X5 | 2.24 | 1 | 2.24 | 1.53 | 0.222 8 |
| X4X5 | 0.034 | 1 | 0.034 | 0.023 | 0.879 8 |
| 残差 Residual | 70.36 | 48 | 1.47 | ||
| 失拟项 Lack of fit | 70.33 | 6 | 11.72 | 17244.59 | < 0.000 1 |
| 纯误差 Pure error | 0.029 | 42 | 6.798E-004 | ||
| 总和 Cor total | 263.72 | 62 |

|
| 图 8 22个配方漆酶酶活差异显著性分析 Figure 8 Analysis of the significant difference in the activity of laccase in 22 formulations 注:图中不同小写字母代表差异性显著,相同字母代表差异性不显著. Note: The different letters represent significant difference, and the same letters represent no significant difference. |
|
|

|
| 图 9 各主料交互作用影响漆酶酶活等高线图 Figure 9 The main interaction effect of the activity of laccase contour map 注:A:小麦秸秆、花生秸秆和油菜秸秆三者交互作用;B:小麦秸秆、大豆秸秆和油菜秸秆三者交互作用;C:小麦秸秆、水稻秸秆和油菜秸秆三者交互作用;D:油菜秸秆、花生秸秆和大豆秸秆三者交互作用;E:油菜秸秆、水稻秸秆、花生秸秆三者交互作用;F:花生秸秆、大豆秸秆和水稻秸秆三者交互作用;G:大豆秸秆、水稻秸秆和小麦秸秆三者交互作用. Note: A: The interaction of wheat straw, peanut straw and rape straw; B: The interaction of wheat straw, soybean straw and rape straw; C: The interaction of wheat straw, rice straw and rape straw; D: The interaction of rape straw, peanut straw and soybean straw; E: The interaction of rape straw, rice straw, peanut straw; F: The interaction of peanut straw, soybean straw and rice straw; G: The interaction of soybean straw, rice straw and wheat straw. |
|
|
| 配方 Formula |
多酚氧化酶酶活 Polyphenol oxidase activity (U/g) |
纤维素酶酶活 Cellulase activity (U/g) |
漆酶酶活 Laccase activity (U/g) |
菌丝生长速率 Mycelium growth rate (mm/d) |
| VF | 1.67±0.013a | 7.07±0.122a | 10.76±0.120a | 6.60±0.070a |
| CK | 1.64±0.026a | 4.07±0.093b | 6.70±0.270b | 6.55±0.127a |
| 注:VF:验证配方;CK:对照配方;a、b:差异显著(P < 0.05). Note: Verification formula; CK: Control check; a, b: Difference was significant (P < 0.05). |
||||
本试验分析了各评价指标之间的相关性,发现红平菇菌丝生长速率与漆酶活性之间无相关性,与贾素巧[26]研究的猴头菌菌丝生长速率与漆酶活性之间无相关性的结论相符。如在本研究的10号和15号配方中,漆酶活性无差异显著性,但是菌丝生长速率却有较大差异。在本研究中我们还可以看到不同栽培基质对红平菇生长过程中胞外酶活影响较大,与吴周斌等[19]研究的不同培养料对真姬菇胞外酶活性影响较大的结果相符。不同基质中木质素、纤维素等含量均有较大差异,说明底物对酶活影响很大。分析22个配方纤维素酶活性,发现大多数配方的纤维素酶活性均高于对照配方,与胡传琪等[27]研究的滑菇菌丝生长时期纤维素酶活性高于木屑基料的结论相符。推测木屑中纤维素含量较秸秆粉少,因此纤维素酶活性比秸秆粉中的低。纤维素是影响纤维素酶活性的主要因素,但本试验中发现有4个配方纤维素酶活性比对照配方低,推测还有其他影响纤维素酶活性的底物。
本研究通过混料设计中的单纯型格子法筛选适合红平菇生长的农业剩余物,试验结果表明:油菜秸秆多酚氧化酶活性最高,花生秸秆与油菜秸秆1׃1交互作用可显著降低多酚氧化酶活性;大豆秸秆基质中多酚氧化酶活性最低,与花生秸秆1׃1交互作用可显著增高多酚氧化酶活性;小麦秸秆纤维素酶活性最高,与花生秸秆秸秆1׃1交互作用可显著降低纤维素酶活性;大豆秸秆纤维素酶活性最低,与花生秸秆1׃1交互作用可显著增加纤维素酶活性;小麦秸秆与花生秸秆1׃1交互作用可显著提高漆酶活性,大豆秸秆与花生秸秆1׃1交互作用可显著提高漆酶活性;油菜秸秆与花生秸秆1׃1交互作用可显著降低菌丝生长速率。由此也可以发现,花生秸秆比较适合做一种调节材料。小麦秸秆和大豆秸秆有利于红平菇菌丝的生长,当大豆秸秆基质中加入少量的小麦秸秆时,可以显著提高菌丝的生长速率。大豆秸秆基质中漆酶活性最高而纤维素酶活性最低;小麦秸秆基质中纤维素酶活性最高而漆酶活性较低。推测大豆秸秆中木质素含量较高、纤维素含量较低,而小麦秸秆中纤维素含量较高、木质素含量较低。
设定各评价指数的期望响应值,最终通过得到一个适合红平菇菌丝体生长的“以草代木”配方:小麦秸秆55.3%、大豆秸秆19.7%、麦麸15.0%、玉米面5.0%、豆粕3.0%、石灰1.0%、石膏1.0%。希望本研究可为以后红平菇代谢基质的研究提供参考依据。
| [1] |
Shi L, Wang YF, Wang JH, et al. Cultivation of Hericium erinaceus by soybean straw[J]. Forest By-Product and Speciality in China, 2015(3): 18-19, 20. (in Chinese) 史磊, 王延锋, 王金贺, 等. 大豆秸秆栽培猴头菇技术[J]. 中国林副特产, 2015(3): 18-19, 20. |
| [2] |
Lin XS. Study on several key technology of Juncao industry development[D]. Fujian: Doctoral Dissertation of Fujian Agricultural and Forestry University, 2013 (in Chinese) 林兴生.菌草产业发展的几个关键技术研究[D].福建: 福建农林大学博士学位论文, 2013 http://cdmd.cnki.com.cn/Article/CDMD-10389-1016190575.htm |
| [3] |
Dai YC, Zhou LW, Yang ZL, et al. A revised checklist of edible fungi in China[J]. Mycosystema, 2010, 29(1): 1-21. (in Chinese) 戴玉成, 周丽伟, 杨祝良, 等. 中国食用菌名录[J]. 菌物学报, 2010, 29(1): 1-21. |
| [4] |
Huang NL. Cultivation of 18 Rare and Delicious Edible Mushrooms[M]. Beijing: China Agricultural Press, 1997: 1-164. (in Chinese) 黄年来. 18种珍稀美味食用菌栽培[M]. 北京: 中国农业出版社, 1997: 1-164. |
| [5] |
Zhang QC, Huang YY, Lai WN, et al. Analysis on the nutritional components of RO-1 of Pleurotus ostreatus[J]. Edible Fungi, 1995(4): 12. (in Chinese) 张其昌, 黄谚谚, 赖万年, 等. 红平菇RO-1营养成分分析[J]. 食用菌, 1995(4): 12. |
| [6] |
Liu ZH. Cultivating technique of liquid isolate and nutrition composition analysis and polysaccharide abstracting technics of Pleurotus djamor[D]. Harbin: Master's Thesis of Northeast Forestry University, 2006 (in Chinese) 刘智会.红平菇液体菌种栽培技术和营养成分分析与多糖提取工艺[D].哈尔滨: 东北林业大学硕士学位论文, 2006 http://cdmd.cnki.com.cn/Article/CDMD-10225-2006110291.htm |
| [7] |
Borges GM, de Barba FFM, Schiebelbein AP, et al. Extracellular polysaccharide production by a strain of Pleurotus djamor isolated in the south of Brazil and antitumor activity on Sarcoma 180[J]. Brazilian Journal of Microbiology, 2014, 44(4): 1059-1605. |
| [8] |
Zhang JJ, Meng GY, Zhang C, et al. The antioxidative effects of acidic-, alkalic-, and enzymatic-extractable mycelium zinc polysaccharides by Pleurotus djamor on liver and kidney of streptozocin-induced diabetic mice[J]. BMC Complementary and Alternative Medicine, 2015, 15: 440. DOI:10.1186/s12906-015-0964-1 |
| [9] |
Men JX, Xing XK, Guo SX. Comparative studies on the cultural characteristics of Armillaria strains associated with sclerotia of Polyporus umbellatus[J]. Mycosystema, 2017, 36(12): 1599-1607. (in Chinese) 门金鑫, 邢晓科, 郭顺星. 伞形多孔菌菌核(猪苓)共生蜜环菌培养特性的比较研究[J]. 菌物学报, 2017, 36(12): 1599-1607. |
| [10] |
Zhang Z. The degeneration of Volvariella volvacea spawn preservation by mycelium subculture[D]. Kunming: Master's Thesis of Kunming University of Science and Technology, 2016 (in Chinese) 张政.草菇菌丝体继代保藏菌种的退化研究[D].昆明: 昆明理工大学硕士学位论文, 2016 http://cdmd.cnki.com.cn/Article/CDMD-10674-1016200331.htm |
| [11] |
Song AR, Guo LZ, Liu ZT, et al. Test and analysis of four kinds of extracellular enzyme activation in fermented liquid of seven white Flammulina velutipes strains[J]. Edible Fungi of China, 1999, 18(4): 31-34. (in Chinese) 宋爱荣, 郭立忠, 刘作亭, 等. 七个白色金针菇菌株发酵液中四种胞外酶活性的测定与分析[J]. 中国食用菌, 1999, 18(4): 31-34. |
| [12] |
Zhuo R, Ma FY, Zhou S, et al. Expression of laccase gene family in different development stages of Pleurotus ostreatus[J]. Mycosystema, 2015, 34(4): 712-716. (in Chinese) 卓睿, 马富英, 周帅, 等. 糙皮侧耳生长发育过程中漆酶基因家族的表达研究[J]. 菌物学报, 2015, 34(4): 712-716. |
| [13] |
Zhao J, Kwan HS. Characterization, molecular cloning, and differential expression analysis of laccase genes from the edible mushroom Lentinula edodes[J]. Applied and Environmental Microbiology, 1999, 65(11): 4908-4913. |
| [14] |
Han ZH, Zhang PQ, Kong XH, et al. Extracellular enzyme activities, mycelial growth rates and fruit body yields of ten Auricularia auricula strains[J]. Acta Edulis Fungi, 2007, 14(4): 41-46. (in Chinese) 韩增华, 张丕奇, 孔祥辉, 等. 黑木耳胞外酶活变化与栽培性状比较的研究[J]. 食用菌学报, 2007, 14(4): 41-46. DOI:10.3969/j.issn.1005-9873.2007.04.007 |
| [15] |
Hu JW, Fan QS, Xiao XN. Study on the functional components in grass-cultivated Ganoderma lucidum and tree-cultivated Ganoderma lucidums[J]. Natural Product Research and Development, 2006, 18(3): 458-460. (in Chinese) 胡居吾, 范青生, 肖小年. 菌草灵芝与段木灵芝的功效成分的比较研究[J]. 天然产物研究与开发, 2006, 18(3): 458-460. DOI:10.3969/j.issn.1001-6880.2006.03.028 |
| [16] |
Feng ZS, Hu QX, Guan DP, et al. Optimization of Pleurotus nebrodensis cultivation substrate using mixture design[J]. Acta Edulis Fungi, 2011, 18(2): 26-29. (in Chinese) 冯作山, 胡清秀, 管道平, 等. 混料设计优化白灵侧耳栽培配方[J]. 食用菌学报, 2011, 18(2): 26-29. DOI:10.3969/j.issn.1005-9873.2011.02.007 |
| [17] |
Liu LQ, Zhang ZJ, Zi HJ, et al. Preliminary study on the cultivation of Pleurotus eryngii with soybean straw[J]. Edible Fungi of China, 2011, 30(2): 33-36. (in Chinese) 刘连强, 张志军, 訾惠君, 等. 大豆秸秆栽培白灵菇配方的初步研究[J]. 中国食用菌, 2011, 30(2): 33-36. DOI:10.3969/j.issn.1003-8310.2011.02.012 |
| [18] |
Zhang P, Xiong BH, Zhao GH, et al. Optimization of Flammulina velutipes cultivation substrate using mixture design[J]. Acta Edulis Fungi, 2016, 23(2): 31-36. (in Chinese) 张鹏, 熊炳辉, 赵光辉, 等. 混料设计优化金针菇栽培配方[J]. 食用菌学报, 2016, 23(2): 31-36. |
| [19] |
Wu ZB, Zhang J, Wang JM, et al. Effect of different cultivation substrates on extracellular enzyme activities of Hypsizygus marmoreus[J]. Northern Horticulture, 2015(8): 145-149. (in Chinese) 吴周斌, 张健, 王佳敏, 等. 不同培养料对真姬菇胞外酶活性的影响[J]. 北方园艺, 2015(8): 145-149. |
| [20] |
An Q, Wu XJ, Wu B, et al. Effects of carbon and nitrogen sources on lignocellulose decomposition enzyme activities in Flammulina velutipes[J]. Mycosystema, 2015, 34(4): 761-771. (in Chinese) 安琪, 吴雪君, 吴冰, 等. 不同碳源和氮源对金针菇降解木质纤维素酶活性的影响[J]. 菌物学报, 2015, 34(4): 761-771. |
| [21] |
Feng GJ, Li SM, Li M, et al. Effect of composts with different C/N on the development of Hericium erinaceus (Bull.) Pers. mycelial and fruitbody[J]. Acta Agriculturae Boreali-Sinica, 2007, 22(S2): 131-135. (in Chinese) 冯改静, 李守勉, 李明, 等. 不同碳氮比栽培料对猴头菌菌丝及子实体生长的影响[J]. 华北农学报, 2007, 22(S2): 131-135. DOI:10.7668/hbnxb.2007.S2.033 |
| [22] |
Niu WJ. Physicochemical composition and energy potential of main crop straw and stalk[D]. Beijing: Doctoral Dissertation of China Agricultural University, 2015 (in Chinese) 牛文娟.主要农作物秸秆组成成分和能源利用潜力[D].北京: 中国农业大学博士学位论文, 2015 http://cdmd.cnki.com.cn/Article/CDMD-10019-1015584263.htm |
| [23] |
Kong QM. Study on near infrared spectrum rapid detection method of crop straw component[D]. Harbin: Doctoral Dissertation of Northeast Agricultural University, 2015 (in Chinese) 孔庆明.大豆秸秆成分近红外光谱分析快速检测方法研究[D].哈尔滨: 东北农业大学博士学位论文, 2015 http://cdmd.cnki.com.cn/Article/CDMD-10224-1015395544.htm |
| [24] |
Ma F. Mechanisim and test research on peanut straw stalk feed molding[D]. Shenyang: Doctoral Dissertation of Shenyang Agricultural University, 2017 (in Chinese) 马方.预处理方式对花生秸秆成型性能影响的试验研究[D].沈阳: 沈阳农业大学博士学位论文, 2017 http://cdmd.cnki.com.cn/Article/CDMD-10157-1017246991.htm |
| [25] |
He PX, Zhang Y, Song YF. Determination of C/N ratio of several kinds of crop straw[J]. Edible Fungi, 2001(4): 15-16. (in Chinese) 何培新, 张炎, 宋永芳. 食用菌原料——几种作物秸秆的碳氮比测定[J]. 食用菌, 2001(4): 15-16. |
| [26] |
Jia SQ. Effects of carbon and nitrogen nutrient on growth development and the extracellular enzyme activity in Hericium erinaceus[D]. Baoding: Master's Thesis of Agricultural University of Hebei, 2006 (in Chinese) 贾素巧.碳氮营养对猴头菌生长发育及胞外酶活性的影响[D].保定: 河北农业大学硕士学位论文, 2006 http://cdmd.cnki.com.cn/Article/CDMD-11920-2006075233.htm |
| [27] |
Hu CQ, Gang J, Chen JH, et al. Research on the cellulase and laccase of Pholiota nameko Zinc-riching cultivated in straw medium[J]. Food and Fermentation Industries, 2015, 41(4): 121-124. (in Chinese) 胡传琪, 冮洁, 陈静慧, 等. 以秸秆为基料富锌栽培滑菇的纤维素酶和漆酶的变化规律[J]. 食品与发酵工业, 2015, 41(4): 121-124. |