 2019, Vol. 46
2019, Vol. 46扩展功能
文章信息
- 武晓桐, 姜欣, 盛思远, 许本姝, 门梦琪, 孟庆欣, 邓利廷, 成利军, 张文浩, 许修宏
- WU Xiao-Tong, JIANG Xin, SHENG Si-Yuan, XU Ben-Shu, MEN Meng-Qi, MENG Qing-Xin, DENG Li-Ting, CHENG Li-Jun, ZHANG Wen-Hao, XU Xiu-Hong
- 静态好氧高温牛粪堆肥中nirK型反硝化细菌群落动态变化
- Dynamics of nirK type denitrifying bacterial community in static aerobic high temperature composting
- 微生物学通报, 2019, 46(5): 986-996
- Microbiology China, 2019, 46(5): 986-996
- DOI: 10.13344/j.microbiol.china.180403
-
文章历史
- 收稿日期: 2018-05-24
- 接受日期: 2018-08-01
- 网络首发日期: 2018-08-09

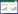




堆肥化作为处理牛粪秸秆最有效的方法之一,已经得到广泛应用。但是传统自然堆肥需要通过翻堆和通风等方法调整堆体温度和溶氧量使堆肥发酵更均匀,不仅增加了能耗,还导致以NH3为主的含氮化合物的挥发,降低了堆肥肥效[1]。另外,由于堆体中温度和氧气分布不均,堆肥过程中会出现堆体表面温度低以及深层厌氧的现象,导致表层和深层堆肥效果不佳。本研究采用静态高温堆肥技术,通过人为控制环境温度,增加堆体与环境的接触表面积,在减少能耗的同时克服堆体表面温度低和深层厌氧等技术缺陷。
在堆肥过程中不同反硝化微生物相互作用产生大量气态氮,导致氮素流失致使肥效降低,同时造成环境污染[2],因此,堆肥化进程中氮素转化规律及控制氮素损失成为研究热点。反硝化作用主要反应过程为NO3−→NO2−→NO→N2O→N2,每一步都由特定的酶催化,依次是硝酸盐还原酶、亚硝酸盐还原酶、一氧化氮还原酶和一氧化二氮还原酶,每一种酶都有对应的编码基因,即4种反硝化功能基因,分别为Nar、Nir、Nor和Nos[3]。亚硝酸盐还原酶(Nir)将NO2−还原成NO,是反硝化过程中第一个可以产生气体的过程,也是反硝化微生物研究过程中用得最多的功能基因产物[4]。功能基因Nir主要分为两种,一种是可溶性含铜酶基因Cu-Nir,即nirK;另一种为细胞色素还原酶基因cd1-Nir,即nirS,虽然这两种基因的功能相同,但是两者酶的结构与催化点位完全不同,往往不能同时出现在一个细胞中[5]。其中,nirK存在于许多亲缘关系较远的菌群中,而含nirS的反硝化细菌主要为假单胞菌[6]。Maeda等[7]利用PCR-DGGE技术研究牛粪堆肥中反硝化细菌群落动态变化,结果表明nirS基因反硝化细菌群落较少,nirK基因编码的亚硝酸还原酶在反硝化过程中具有决定性的作用。近年来nirK基因作为分子标记被广泛应用于不同生境中反硝化细菌群落结构的研究,例如森林土壤、河底沉积物和耕地土壤等[8-10]。因此,将nirK功能基因作为分子标记可以真实反映堆肥环境中反硝化菌群结构。
本研究以nirK功能基因为标记,通过高通量测序技术分析牛粪秸秆堆肥过程中反硝化细菌的群落结构和多样性变化,揭示新型静态高温堆肥过程中反硝化细菌群落组成的动态变化,应用Spearman相关性热图分析堆肥中优势反硝化细菌类别与理化指标之间的相关性。
1 材料与方法 1.1 主要试剂和仪器DNA提取试剂盒,Omega公司;凝胶回收试剂盒,Axygen公司;QuantiFluor™-ST蓝色荧光定量系统,Promega公司。
连续流动分析仪,水尔分析仪器(上海)有限公司;PCR仪,Thermo Fisher Scientific公司。
本试验装置的截面如图 1所示。装置是由铁丝网编织成的同轴圆筒,距离地面高20 cm。其内筒直径60 cm,外筒直径180 cm,高为160 cm。堆肥原料填充在图 1中的灰色区域,形成一个中空的圆柱体(可以容纳3.62 m3堆肥原料),利于堆肥原料的内表面和外表面都能接触空气,保证堆体内的氧气充足。将该装置置于发酵仓内,发酵仓内温度保持在40 ℃左右并定期通风,保持仓内空气新鲜。

|
| 图 1 堆肥装置的横截面 Figure 1 Cross section of the composting system |
|
|
实验堆肥材料为牛粪和水稻秸秆,原料取自黑龙江省哈尔滨市阿城区畜牧业基地,主要理化参数见表 1。将水稻秸秆切成3 cm−5 cm的小段与粉碎的牛粪按质量比1:3.5充分混合,堆肥初始C/N约为35:1。堆肥化过程采用静态好氧高温堆肥,全程共17 d,分别在第0、2、4、6、8、10、12、14、17天时取样。取样方法为:分别从装置的上层(距表面10 cm−15 cm,A1和A2点)、中层(距表面80 cm−85 cm,A3和A4点)、下层(距表面145 cm−150 cm,A5和A6点)三层取样(图 1),每个位置都均匀地在同层取4个点,所有点的样品充分混匀后为这个时期的样品,每个样品取1 kg。另外,第0天(初始期)、第2天(升温期)、第6天(高温期)、第12天(高温期后期)和第17天(腐熟期)混合后的样品置于−80 ℃环境下保存,用于后续分子实验分析,并分别命名为C0、C2、C6、C12和C17样品。
| 堆肥材料 Compost material |
牛粪 Dairy manure |
水稻秸秆 Rice stalk |
| 含水率WC (%) | 48.26±1.34 | 11.93±0.45 |
| 总有机碳TOC (%) | 33.68±0.96 | 43.52±1.07 |
| 凯氏氮TKN (%) | 1.89±0.16 | 0.88±0.09 |
| 碳氮比C/N | 17.82±0.82 | 49.45±3.17 |
| 注:WC:含水率;TOC:总有机碳;TKN:凯氏氮. Note: WC: Water content; TOC: Total organic carbon; TKN: Total kjeldahl nitrogen. |
||
在堆体不同深度(距离堆体外表面10 cm和30 cm)的上、中、下三层(距离堆体底部15、80、145 cm)的6个不同位置,每天使用精密温度计分别测其温度并取外层(距离堆体外表面10 cm)及中心(距离堆体外表面30 cm)不同层温度的平均值,同时记录上、中、下三层的环境温度。含水率(Water content,WC)的测定采用恒重法。pH的测定是将堆肥样品按1:10 (质量浓度比)比例加入去离子水,振荡过滤后的滤液用数字pH仪测定。总有机碳(Total organic carbon,TOC)和凯氏氮(Total kjeldahl nitrogen,TKN)分别采用重铬酸钾容量法和凯氏定氮法[11]测定,两者的比值即为碳氮比(C/N)。水溶性铵态氮(NH4+-N)和硝态氮(NO3−-N)的含量采用连续流动分析仪测定。种子发芽指数(Germination index,GI)使用独行菜(Lepidium sativum L.)种子[12]测定。将堆肥样品按1:7.5 (质量浓度比)比例加入去离子水,取5 mL提取液滴入含有滤纸的培养皿中,并将20粒种子均匀分布在滤纸上,在25 ℃黑暗条件下培养48 h。根据计算公式:GI (%)=处理平均发芽率×处理平均根长/(对照平均发芽率×对照平均根长)×100,计算种子发芽指数。以上除温度外的其他指标,每个样品均进行3次重复测定。
1.4 样品总DNA提取使用Omega的Soil DNA Isolation Kit提取堆肥样品总DNA,利用1%琼脂糖凝胶电泳检测基因组DNA的完整性,利用NanoDrop 2000检测基因组DNA的浓度和纯度,保存在−20 ℃待用。
1.5 nirK型反硝化细菌高通量测序分别以各样品总DNA为模板扩增nirK基因。引物为nirKlaCuF (5′-ATCATGGTSCTGCCGCG-3′)和nirKR3CuR (5′-GCCTCGATCAGRTTGTGGTT-3′)[13]。PCR扩增体系(20 μL):5×FastPfu buffer 4 μL,dNTPs (2.5 mmol/L) 2 μL,DNA 10 ng,BSA 0.2 μL,正、反向引物(5 µmol/L)各0.8 µL,FastPfu DNA polymerase (2.5 U/μL) 0.4 μL,补足ddH2O至20 μL。PCR反应条件:95 ℃ 3 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 45 s,35个循环;72 ℃ 10 min;4 ℃停止反应。PCR产物用2%琼脂糖凝胶电泳检测,切胶使用AxyPrepDNA凝胶回收试剂盒回收PCR产物,Tris-HCl洗脱,再用2%琼脂糖电泳检测。参照电泳初步定量结果,将PCR产物用QuantiFluorTM-ST蓝色荧光定量系统进行定量检测,按照各个样本的测序量要求进行相应比例的混合。最后进行MiSeq测序。原始数据提交至NCBI数据库,登录号为SRP136699。
1.6 生物信息处理测序的原始数据经过Fast QC软件进行质量控制,PRINSEQ软件过滤小于250 bp序列,得到优化序列。将相似性大于97%的优化序列划分为一个操作分类单元(Operational taxonomic unit,OTU);基于OTU计算覆盖度(Coverage)和Shannon多样性指数;以R语言统计物种并绘制Venn图;利用样本间距离Heatmap分析其OTU的相似性;对比GenBank的功能基因数据库(Release7.3,http://fungene.cme.msu.edu/),采用RDP分类器(Version 2.2 Release 11.1,http://rdp.cme.msu.edu/)和贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析,并在各个分类水平上统计每个样品的群落组成。利用Spearman相关性矩阵分析堆肥样品的理化指标与nirK型反硝化细菌群落的关系。
1.7 数据处理基础数据利用Excel 2013进行分析,采用SPSS 22.0对堆肥不同时期理化参数进行单因素方差分析(One-way ANOVA,Tukey HSD法)。
2 结果与分析 2.1 堆肥过程中理化参数及GI指标的变化堆肥过程中,微生物快速分解有机质,产生CO2和水,并释放出大量的热,使堆肥物料的温度上升。堆肥过程中温度的变化可反映微生物活性的变化和有机质的降解程度,并影响最终堆肥产品的成熟度和质量[14]。图 2表示堆肥过程中温度的变化,从中可以看出,外层和中心温度在堆肥开始后均迅速升高,在第3天分别达到55.8 ℃和59.8 ℃;然后温度保持上升趋势并都在第6天达到最高温度,分别为56.9 ℃和61.9 ℃。外层和中心的温度从第3天直到第12天均保持在55 ℃以上,在第12天后,温度呈缓慢降低趋势。堆肥过程中温度的变化表明了3个阶段:升温期(第0−2天)、高温期(第3−12天,大于55 ℃)、降温及腐熟期(第13−17天)。根据国家粪便无害化卫生标准(GB7959-87)规定,堆肥过程的温度必须达50−55 ℃,并持续5−7 d。本研究中外层和中心温度连续9 d保持在55 ℃以上,所以无论堆肥物料的外层还是中心部分,其温度均符合标准。

|
| 图 2 堆肥过程中温度的变化 Figure 2 Changes of temperature during composting process |
|
|
表 2是其他理化参数的测定结果。从表 2可以看出,在堆肥过程中含水率整体呈下降趋势,腐熟期达到51.0%;pH整体呈下降的趋势,其变化范围为8.32−8.77,一直处于弱碱性的状态,有利于微生物保持良好的活性[15]。碳氮比(C/N)是评价堆肥产品腐熟程度的一项重要指标,在表 2中C/N从初始值35.0下降到20.0 (P < 0.05),整体呈下降趋势。当C/N≤20.0时,认定堆肥已经达到腐熟[16]。种子发芽指数(GI)是指示堆肥腐熟程度的生物学指标[17],当GI≥80.0%时表示堆肥产品已经达到腐熟,其植物毒性已经丧失[17-18]。从表 2可以看到,GI整体呈上升的趋势,从堆肥前期的37.63%上升到97.70% (P < 0.05),表明堆肥产品符合腐熟标准。随着堆肥的进程,铵态氮含量变化呈先升后降的趋势。在第0−2天,铵态氮含量从864.8 mg/kg上升至932.2 mg/kg (P < 0.05),从第4天开始,铵态氮含量呈显著下降趋势(P < 0.05),并在堆肥结束后达到最低,为311.3 mg/kg。堆肥过程中硝态氮含量整体呈上升趋势,由初始阶段的55.8 mg/kg上升至堆肥结束后的95.5 mg/kg (P < 0.05)。可能是由于过高的温度抑制了硝化微生物的活性[19-20],使硝态氮生成和积累的速度缓慢。根据以上所有指标综合判断,利用堆肥装置来处理牛粪和水稻秸秆的混合物,在堆肥第17天时达到腐熟要求。
| 样品 Samples |
pH | 含水率 WC (%) |
铵态氮 NH4+-N (mg/kg) |
硝态氮 NO3–-N (mg/kg) |
碳氮比 C/N |
种子发芽指数 GI (%) |
| 0 | 8.77±0.09a | 72.0±1.6a | 864.8±11.4b | 55.8±1.4f | 35.0±0.5a | 37.63±1.78f |
| 2 | 8.70±0.03a | 71.0±0.9a | 932.2±12.7a | 57.9±1.0f | 32.2±1.3a | 40.25±1.98f |
| 4 | 8.68±0.07a | 69.5±0.7ab | 881.0±8.5b | 62.6±1.0e | 28.9±1.2b | 50.74±1.78e |
| 6 | 8.67±0.05ab | 67.0±0.7b | 774.1±2.4c | 71.9±1.3d | 25.7±0.8c | 54.68±2.05e |
| 8 | 8.62±0.09ab | 63.4±2.2c | 658.1±5.6d | 81.3±0.5c | 24.5±1.1cd | 77.14±2.44d |
| 10 | 8.59±0.02abc | 59.0±1.0d | 553.0±15.3e | 85.1±1.4bc | 23.5±0.6cd | 85.85±1.22c |
| 12 | 8.48±0.01bcd | 55.0±0.6e | 466.2±4.5f | 88.7±1.5b | 23.1±0.9cde | 89.04±1.93bc |
| 14 | 8.42±0.06cd | 52.0±1.0ef | 388.4±4.8g | 89.7±0.6b | 21.7±1.0de | 92.84±1.33ab |
| 17 | 8.32±0.01d | 51.0±0.5f | 311.3±2.5h | 95.5±1.2a | 20.0±0.4e | 97.70±1.95a |
| 注:字母不同者差异显著(P < 0.05). Note: Different superscripts denote significant differences (P < 0.05). |
||||||
所有堆肥样品通过高通量测序共获得142 417条高质量序列,依次为28 199、43 040、19 399、19 581和32 198条。以97%相似度划分,共得到212个OTU。从图 3中可以看出,在堆肥过程中5个样品检测到的OTU数量先逐渐减少,初始期样品(C0)中最多为206,在高温期后期样品(C12)中达到最小值为146,腐熟期样品(C17)中的OTU数量为164个,说明本次堆肥中在堆肥初期的反硝化细菌群落丰富度最高,随着堆肥的进行,反硝化细菌群落丰富度逐渐下降,在高温期末期达到最低,随后又升高。5个样品中共96个相同的OTU,占总数的39.67%,其中C0中有7个特有OTU,C17中有12个特有OTU,C2、C6和C12没有特有OTU,说明在升温期和高温期堆肥中反硝化细菌的种类相似。在表 3中各个样品文库的覆盖率均超过99.7%,说明测序结果能够代表堆肥中nirK型反硝化细菌群落的真实情况。

|
| 图 3 不同堆肥时期反硝化细菌群落韦恩图分析 Figure 3 Venn diagram of the denitrifier community in the different periods of composting 注:Cn代表堆肥第n天. Note: Cn represents nth day in composting. |
|
|
| 样品 Samples |
优化序列 Optimal sequence |
OTUs | 覆盖率 Coverage |
Shannon指数 Shannon index |
| C0 | 28 199 | 206 | 0.999 | 3.32 |
| C2 | 43 040 | 200 | 0.997 | 2.47 |
| C6 | 19 399 | 175 | 0.999 | 2.44 |
| C12 | 19 581 | 146 | 0.997 | 2.06 |
| C17 | 32 198 | 164 | 0.998 | 1.86 |
| Total | 142 417 |
堆肥过程中nirK型反硝化细菌α多样性统计结果见表 3。Shannon指数是用来估算样品中微生物多样性的指数之一,从表 3可知Shannon指数由大到小依次为C0 > C2 > C6 > C12 > C17,即堆肥初始期nirK型反硝化细菌群落多样性最高,降温期和腐熟期较低。
根据5个堆肥样品中OTU的组成进行样品之间距离Heatmap分析(图 4),在图 4中样本之间的距离用一定颜色梯度进行表示(图中右侧为颜色梯度代表的数值),颜色越深,样本之间距离越大,表示细菌群落结构差异性越大。从图 4中分析得出,C12和C17之间的距离最近,为0.334 2,即两者OTU组成相近;C6与C12之间的距离次之,为0.407 4,即两者OTU组成也相似;C0与C2的OTU组成也相近,为0.431,但是差异性大于C12与C17,即两者之间的反硝化细菌群落结构差异性大于C12和C17的,说明在堆肥后期反硝化细菌群落结构趋于稳定;高温期C6与其他不同时期之间的距离从大到小依次为C0 > C2 > C17 > C12,即nirK型反硝化细菌群落结构在堆肥高温期前后差异变化显著。

|
| 图 4 堆肥样品在OTU水平距离Heatmap Figure 4 Distances heatmap on OTUs level of composting sample |
|
|
在5个堆肥样品中的212条OTU分属于3门5纲8目15科17属20种。图 5A为门分类水平的nirK型反硝化细菌组成变化(将相对丰度 < 0.1%的部分合并为Others在图中显示),主要为变形菌门(Proteobacteria)和未分类门的反硝化细菌(Unclassified_k_norank_d_Bacteria),在不同时期堆肥样品C0、C2、C6、C12和C17中变形菌门所占比例分别为78.6%、71.1%、35.8%、41.3%和56.3%,变化趋势为先减少后增加,在堆肥前期相对丰度最高,高温期(C6期)最少;C0、C2、C6、C12和C17中未分类门的反硝化细菌所占比例分别为21.1%、28.8%、64.0%、58.7%和43.6%,变化趋势为先增多后减少,在高温期(C6期)相对丰度达到最高。

|
| 图 5 堆肥样品中nirK型反硝化细菌在门(A)及目(B)分类水平上群落组成的动态变化 Figure 5 Dynamic changes of community composition of nirK type denitrifier community during composting process at the level of phylum (A) and order (B) |
|
|
图 5B为目分类水平的nirK型反硝化细菌分类(将相对丰度 < 1%的部分合并为Others在图中显示),主要包括:未分类门的反硝化细菌(Unclassified_k_ norank_d_Bacteria)、伯克氏菌目(Burkholderiales)、根瘤菌目(Rhizobiales)、未分类纲的变形菌门的反硝化细菌(Unclassified_p_Proteobacteria)和红杆菌目(Rhodobacterales)。在C0中主要含有Burkholderiales (42.2%)、Rhizobiales (8.2%)、Unclassified_p_ Proteobacteria (26.7%)和Rhodobacterales (1.2%)。堆肥升温期C2中主要含有Burkholderiales (47.9%)、Rhizobiales (5.2%)、Unclassified_p_Proteobacteria(17.4%)和Rhodobacterales (0.3%);堆肥高温期C6中主要含有Burkholderiales (11.9%)、Rhizobiales (14.1%)、Unclassified_p_Proteobacteria (9.5%)和Rhodobacterales (0.19%);堆肥高温期末期C12中主要含有Burkholderiales (10.1%)、Rhizobiales (24.6%)和Unclassified_p_Proteobacteria (6.5%);堆肥降温期和腐熟期C17中主要含有Burkholderiales (19.6%)、Rhizobiales (33.9%)和Unclassified_p_Proteobacteria (2.7%)。Burkholderiales、Rhizobiales、Unclassified_p_ Proteobacteria和Rhodobacterales都是变形菌门。其中Burkholderiales属于β-变形菌纲,Rhizobiales和Rhodobacterales属于α-变形菌纲。在图 5B中基于目分类水平可以看出在堆肥不同时期nirK型反硝化细菌种类类似,但是相对丰度存在差异,Burkholderiales在堆肥初始期和升温期相对丰度较大,并且是这两个时期的优势种群。随着温度的升高,其相对丰度逐渐降低,进入腐熟期后又略微上升。Rhizobiales随着堆肥的进行相对丰度整体呈逐渐上升的趋势,在腐熟期达到最高值。Unclassified_p_Proteobacteria的相对丰度变化趋势则相反,在堆肥初始期相对丰度最高,随着温度的升高逐渐减少,在腐熟期达到最低。Rhodobacterales主要存在于堆肥初始期,随着堆体温度上升相对丰度逐渐减少,在高温期达到0.19%,高温期末期其相对分度更是低于0.01%。
2.4 堆肥中nirK型反硝化细菌群落组成与理化指标的相关关系探讨Spearman相关性热图(Heatmap)表示的是微生物分类与环境变量比较,评估微生物分类与环境变量之间的相关性,基本输出的是一个距离矩阵,表示群落中每个微生物与每个环境因子变量之间的相关系数。在图 6中,Unclassified_k_norank_d_Bacteria与TOC、C/N、WC和pH之间呈显著负相关(P < 0.05),与TKN和NO3−-N之间呈显著正相关(P < 0.05)。Unclassified_o_Burkholderiales只与NH4+-N之间呈显著正相关(P < 0.05)。Unclassified_o_Rhizobiales与NH4+-N之间呈极显著负相关(P < 0.001),与WC、C/N、TOC和pH之间呈显著负相关(P < 0.05),与TKN和NO3−-N之间呈显著正相关(P < 0.05)。Unclassified_p_Proteobacteria与NH4+-N之间呈极显著正相关(P < 0.001),与TKN和NO3−-N之间呈显著负相关(P < 0.05),与WC、C/N、TOC和pH之间呈显著正相关(P < 0.05)。产碱杆菌科的Pusillimonas属与NH4+-N之间呈极显著正相关(P < 0.001),与TKN和NO3−-N之间呈显著负相关(P < 0.05),与WC、C/N、TOC和pH之间呈显著正相关(P < 0.05)。副球菌属(Paracoccus)与TKN和NO3−-N之间呈极显著负相关(P < 0.001),与WC、C/N、TOC和pH之间呈极显著正相关(P < 0.001),与NH4+-N之间呈显著正相关(P < 0.05)。所有优势菌属与温度没有相关性。

|
| 图 6 Spearman相关性热图 Figure 6 Spearman correlation heatmap 注:热图右侧色卡是不同R值的颜色分区,蓝色代表正相关,0 < R < 1;白色代表负相关,−1 < R < 0. *:P < 0.05;**:P < 0.01;***:P < 0.001. Note: The color card on the right side of the heatmap is a color partition with different R values. The blue color represents positive correlation, 0 < R < 1, and white represents negative correlation, −1 < R < 0. Ammonium nitrogen: NH4+-N; Nitrate nitrogen: NO3–-N. *: P < 0.05; **: P < 0.01; ***: P < 0.001. |
|
|
在堆肥过程中,随着温度的升高,堆体中微生物的活性也会增强[21]。据文献[16, 22-25]报道在高温期,嗜热微生物在有机底物的降解中充当重要角色,能够降解超过2/3的木质素,而嗜温微生物主要降解易降解的有机底物,说明在堆肥过程中维持较长的高温期可以促进有机底物的降解,促进堆肥的进程,缩短堆肥周期。本研究中堆肥全程共17 d,堆体温度高于40 ℃以上就达到了16 d,在此高温条件下微生物的代谢加快,降解有机物效率提高,为缩短堆肥周期创造了良好的环境条件。通过分析各项堆肥理化参数以及生物学指标发现堆肥已经基本腐熟,相比传统的一般为期40−63 d的自然堆肥周期[22, 26],静态好氧高温堆肥技术缩短了堆肥周期。
利用高通量测序技术分析牛粪和水稻秸秆混合堆肥过程中nirK型反硝化细菌群落动态变化,各样品的覆盖率和稀释曲线说明测序结果能够代表堆肥中nirK型反硝化细菌群落的真实情况。韦恩图分析表明在堆肥各个时期nirK型反硝化细菌群落丰富度变化依次为C0 > C2 > C6 > C17 > C12。α多样性分析表明堆肥前期nirK型反硝化细菌群落多样性最高,堆肥后期较低,Shannon指数变化范围为1.86−3.32。β多样性分析结果表明堆肥不同时期样本间物种的丰度分布差异显著,经过高温期后堆肥中nirK型反硝化细菌差异变化显著。陆彦宇利用牛粪和中药渣进行堆肥发现细菌在堆肥高温期前后种群发生了很大的变化[27],这与本研究结果相符。
针对各种环境样品的研究表明,最常见的反硝化细菌都属于变形菌门,近年来国内外有关农业废弃物堆肥中反硝化微生物种类的报道也主要为变形菌门[28],本研究分别从门和目分类水平上对牛粪和水稻秸秆混合堆肥中nirK型反硝化细菌群落组成进行了分析,结果表明在堆肥中反硝化细菌主要为变形菌门;另外,未分类反硝化细菌占比也比较大,表明在堆肥中存在大量未知的反硝化细菌。康鹏亮等[29]利用高通量测序技术分析了西安市典型景观水体中反硝化细菌种群结构,发现未分类反硝化细菌的占比较大,这与本研究结果相符。在污水处理中nirK型反硝化细菌多是α-变形菌纲[30-31],在稻田中nirK型反硝化细菌多是α-变形菌纲根瘤菌目[8, 32-33],在本研究中α-变形菌纲的nirK型反硝化细菌占比也较大。目分类水平nirK型反硝化细菌群落结构分析表明,堆肥中反硝化细菌主要是α-变形菌纲下的根瘤菌目和红杆菌目,以及β-变形菌纲下的伯克氏菌目。周唯采用PCR-DGGE技术分析农业废弃物好氧堆肥中反硝化细菌的多样性,发现在农业废弃物堆肥过程中nirK基因主要集中于变形菌门,其中包括α-变形菌纲根瘤菌目中叶杆菌科、慢性根瘤菌科、根瘤菌科和布鲁氏菌科,红杆菌目中红杆菌科,以及β-变形菌纲伯克氏菌目中产碱杆菌科[28],与本研究结果相符。
Spearman相关性分析表明,除了未分类属产碱杆菌科的反硝化细菌(Unclassified_f_Alcaligenaceae)和苍白杆菌属(Ochrobactrum)外,其余10个nirK型反硝化细菌菌属变化与环境因子相关,说明在堆肥过程中环境因素与反硝化细菌群落结构变化相关[34]。在本研究中外部温度控制在40 ℃以上,使堆肥整个过程中温度的变化不是很明显,特别是在堆肥后期基本维持在40 ℃左右,因此nirK型反硝化细菌群落结构变化与温度之间不相关。除了Unclassified_k_ norank_d_Bacteria和Unclassified_o_Rhizobiales之外,剩余8个菌属都与TOC、C/N、WC、pH呈正相关,与TKN、NO3−-N呈负相关,而Unclassified_k_ norank_d_Bacteria和Unclassified_o_Rhizobiales正好相反。胡春晓等[2]利用定量聚合酶链式反应技术分析农业废弃物好氧堆肥环境因子对nirK、nirS和nosZ数量的影响,结果表明nirK型反硝化细菌与pH显著正相关,与含水率有一定相关性,但不显著;与NO3−-N没有相关性。含水率作为堆肥中微生物生理活动吸收营养物质重要的溶解和传输介质,对微生物活性有很大影响[35]。有研究表明如果堆肥中含水率过高,堆体中的溶氧量就会减少,可能会形成局部厌氧环境,更有利于反硝化细菌生长繁殖,因为大多数反硝化细菌为厌氧微生物[2, 34]。NO3−-N作为反硝化过程的电子受体,对反硝化细菌群落变化有重要影响[36-37]。马丽红等[38]对牛粪堆肥过程中各类氮素形态物质与微生物群落结构变化的关系作了分析,结果表明反硝化细菌数量与NO3−-N和全氮含量呈负相关,与本研究结果相符。
研究表明,静态好氧高温堆肥技术可以缩短堆肥周期。在堆肥过程中,nirK型反硝化细菌群落结构变化显著且受到堆肥理化因子的影响。本研究为丰富堆肥氮素循环理论和改进堆肥工艺流程提供了一定的科学依据。
| [1] |
Szanto GL, Hamelers HVM, Rulkens WH, et al. NH3, N2O and CH4 emissions during passively aerated composting of straw-rich pig manure[J]. Bioresource Technology, 2007, 98(14): 2659-2670. DOI:10.1016/j.biortech.2006.09.021 |
| [2] |
Hu CX, Chen YN, Zhang JC, et al. Effects of physico-chemical parameters on the abundance of the denitrification-associated genes nirK, nirS and nosZ during agricultural waste composting[J]. Environmental Science, 2013, 34(3): 1196-1203. (in Chinese) 胡春晓, 陈耀宁, 张嘉超, 等. 农业废物好氧堆肥中环境因子对nirK、nirS和nosZ数量的影响[J]. 环境科学, 2013, 34(3): 1196-1203. |
| [3] |
Borrerode-de Acu a JM, Rohde M, Wissing J, et al. Protein network of the Pseudomonas aeruginosa denitrification apparatus[J]. Journal of Bacteriology, 2016, 198(9): 1401-1413. DOI:10.1128/JB.00055-16 |
| [4] |
Kandeler E, Deiglmayr K, Tscherko D, et al. Abundance of narG, nirS, nirK, and nosZ genes of denitrifying bacteria during primary successions of a glacier foreland[J]. Applied and Environmental Microbiology, 2006, 72(9): 5957-5962. DOI:10.1128/AEM.00439-06 |
| [5] |
Zhang SB, He XJ, Wu HL, et al. Advances in molecular ecology technologies for the study of denitrifier communities[J]. Environmental Science & Technology, 2017, 40(6): 120-126. (in Chinese) 张盛博, 何小娟, 吴海露, 等. 分子生态学技术用于反硝化微生物群落研究的进展[J]. 环境科学与技术, 2017, 40(6): 120-126. |
| [6] |
Braker G, Zhou JZ, Wu LY, et al. Nitrite reductase genes (nirK and nirS) as functional markers to investigate diversity of denitrifying bacteria in pacific northwest marine sediment communities[J]. Applied and Environmental Microbiology, 2000, 66(5): 2096-2104. DOI:10.1128/AEM.66.5.2096-2104.2000 |
| [7] |
Maeda K, Morioka R, Hanajima D, et al. The impact of using mature compost on nitrous oxide emission and the denitrifier community in the cattle manure composting process[J]. Microbial Ecology, 2010, 59(1): 25-36. |
| [8] |
Yoshida M, Ishii S, Otsuka S, et al. Temporal shifts in diversity and quantity of nirS and nirK in a rice paddy field soil[J]. Soil Biology and Biochemistry, 2009, 41(10): 2044-2051. DOI:10.1016/j.soilbio.2009.07.012 |
| [9] |
Keil D, Meyer A, Berner D, et al. Influence of land-use intensity on the spatial distribution of N-cycling microorganisms in grassland soils[J]. FEMS Microbiology Ecology, 2011, 77(1): 95-106. |
| [10] |
Wang YY, Lu SE, Chen XM, et al. Analyzing the nitrate reductase gene (nirK) community in the peat soil of the Zoige Wetland of the Tibetan Plateau[J]. Acta Ecologica Sinica, 2017, 37(19): 6607-6615. (in Chinese) 王蓥燕, 卢圣鄂, 陈小敏, 等. 若尔盖高原湿地泥炭沼泽土亚硝酸盐还原酶(nirK)反硝化细菌群落结构分析[J]. 生态学报, 2017, 37(19): 6607-6615. |
| [11] |
Bao SD. Soil and Agricultural Chemistry Analysis[M]. 3rd ed. Beijing: China Agriculture Press, 2000: 39-89. (in Chinese) 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000: 39-89. |
| [12] |
Zucconi F, Pera A, Forte M, et al. Evaluating toxicity of immature compost[J]. Biocycle, 1981, 22(2): 54-57. |
| [13] |
Palmer K, Biasi C, Horn MA. Contrasting denitrifier communities relate to contrasting N2O emission patterns from acidic peat soils in arctic tundra[J]. The ISME Journal, 2012, 6(5): 1058-1077. DOI:10.1038/ismej.2011.172 |
| [14] |
Ren GM, Xu XH, Qu JJ, et al. Evaluation of microbial population dynamics in the co-composting of cow manure and rice straw using high throughput sequencing analysis[J]. World Journal of Microbiology and Biotechnology, 2016, 32(6): 101. DOI:10.1007/s11274-016-2059-7 |
| [15] |
Masó MA, Blasi AB. Evaluation of composting as a strategy for managing organic wastes from a municipal market in Nicaragua[J]. Bioresource Technology, 2008, 99(11): 5120-5124. DOI:10.1016/j.biortech.2007.09.083 |
| [16] |
Xiao Y, Zeng GM, Yang ZH, et al. Continuous thermophilic composting (CTC) for rapid biodegradation and maturation of organic municipal solid waste[J]. Bioresource Technology, 2009, 100(20): 4807-4813. DOI:10.1016/j.biortech.2009.05.013 |
| [17] |
Wang X, Selvam A, Chan MT, et al. Nitrogen conservation and acidity control during food wastes composting through struvite formation[J]. Bioresource Technology, 2013, 147(8): 17-22. |
| [18] |
Paredes C, Cegarra J, Bernal MP, et al. Influence of olive mill wastewater in composting and impact of the compost on a Swiss chard crop and soil properties[J]. Environment International, 2005, 31(2): 305-312. DOI:10.1016/j.envint.2004.10.007 |
| [19] |
Bernal MP, Alburquerque JA, Moral R, et al. Composting of animal manures and chemical criteria for compost maturity assessment. A review[J]. Bioresource Technology, 2009, 100(22): 5444-5453. DOI:10.1016/j.biortech.2008.11.027 |
| [20] |
Tiquia SM, Tam NFY. Characterization and composting of poultry litter in forced-aeration piles[J]. Process Biochemistry, 2002, 37(8): 869-880. DOI:10.1016/S0032-9592(01)00274-6 |
| [21] |
Tiquia SM. Microbial community dynamics in manure composts based on 16S and 18S rDNA T-RFLP profiles[J]. Environmental Technology, 2005, 26(10): 1101-1114. DOI:10.1080/09593332608618482 |
| [22] |
Liu DY, Zhang RF, Wu HH, et al. Changes in biochemical and microbiological parameters during the period of rapid composting of dairy manure with rice chaff[J]. Bioresource Technology, 2011, 102(19): 9040-9049. DOI:10.1016/j.biortech.2011.07.052 |
| [23] |
Mayende L, Wilhelmi BS, Pletschke BI. Cellulases (CMCases) and polyphenol oxidases from thermophilic Bacillus spp. isolated from compost[J]. Soil Biology and Biochemistry, 2006, 38(9): 2963-2966. DOI:10.1016/j.soilbio.2006.03.019 |
| [24] |
Wang CM, Shyu CL, Ho SP, et al. Species diversity and substrate utilization patterns of thermophilic bacterial communities in hot aerobic poultry and cattle manure composts[J]. Microbial Ecology, 2007, 54(1): 1-9. |
| [25] |
Yu HY, Zeng GM, Huang HL, et al. Microbial community succession and lignocellulose degradation during agricultural waste composting[J]. Biodegradation, 2007, 18(6): 793-802. DOI:10.1007/s10532-007-9108-8 |
| [26] |
Zhang J, Sun ZY, Zhong XZ, et al. Maturity and microbial community structure dynamics during simulated windrow composting of solid fraction of dairy manure[J]. Chinese Journal of Applied and Environmental Biology, 2016, 22(3): 423-429. (in Chinese) 张晶, 孙照勇, 钟小忠, 等. 奶牛粪条垛式模拟堆肥腐熟度及微生物群落结构变化[J]. 应用与环境生物学报, 2016, 22(3): 423-429. |
| [27] |
Lu YY. Study on the properties of dissolved organic matter and bacterial structure during composting of dairy manure and chinese herb residues[D]. Nanning: Master's Thesis of Guangxi University, 2017 (in Chinese) 陆彦宇.牛粪和中药渣堆肥化过程中水溶有机质特性及细菌结构的研究[D].南宁: 广西大学硕士学位论文, 2017 http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y3276815 |
| [28] |
Zhou W. Nitrite reductase genes as functional markers to investigate diversity of denitrifying bacteria during agricultural waste composting[D]. Changsha: Master's Thesis of Hunan University, 2014 (in Chinese) 周唯.农业废物好氧堆肥中反硝化细菌的多样性研究[D].长沙: 湖南大学硕士学位论文, 2014 http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2605497 |
| [29] |
Kang PL, Huang TL, Zhang HH, et al. Water quality and diversity of denitrifier community structure of typical scenic water bodies in Xi'an[J]. Environmental Science, 2017, 38(12): 5174-5183. (in Chinese) 康鹏亮, 黄廷林, 张海涵, 等. 西安市典型景观水体水质及反硝化细菌种群结构[J]. 环境科学, 2017, 38(12): 5174-5183. |
| [30] |
Heylen K, Gevers D, Vanparys B, et al. The incidence of nirS and nirK and their genetic heterogeneity in cultivated denitrifiers[J]. Environmental Microbiology, 2006, 8(11): 2012-2021. DOI:10.1111/emi.2006.8.issue-11 |
| [31] |
Heylen K, Vanparys B, Gevers D, et al. Nitric oxide reductase (norB) gene sequence analysis reveals discrepancies with nitrite reductase (nir) gene phylogeny in cultivated denitrifiers[J]. Environmental Microbiology, 2007, 9(4): 1072-1077. DOI:10.1111/emi.2007.9.issue-4 |
| [32] |
Saito T, Ishii S, Otsuka S, et al. Identification of novel betaproteobacteria in a succinate-assimilating population in denitrifying rice paddy soil by using stable isotope probing[J]. Microbes and Environments, 2008, 23(3): 192-200. DOI:10.1264/jsme2.23.192 |
| [33] |
Tsuneda S, Miyauchi R, Ohno T, et al. Characterization of denitrifying polyphosphate-accumulating organisms in activated sludge based on nitrite reductase gene[J]. Journal of Bioscience and Bioengineering, 2005, 99(4): 403-407. DOI:10.1263/jbb.99.403 |
| [34] |
Zhu LP, Xu XH, Zhang WH, et al. Correlationship between denitrifying bacteria and physicochemical factors in cow manure composting[J]. Journal of Agro-Environment Science, 2016, 35(9): 1781-1787. (in Chinese) 朱丽平, 许修宏, 张文浩, 等. 牛粪堆肥中反硝化细菌与理化参数的关系[J]. 农业环境科学学报, 2016, 35(9): 1781-1787. |
| [35] |
Liang C, Das KC, McClendon RW. The influence of temperature and moisture contents regimes on the aerobic microbial activity of a biosolids composting blend[J]. Bioresource Technology, 2003, 86(2): 131-137. DOI:10.1016/S0960-8524(02)00153-0 |
| [36] |
Liu XD, Tiquia SM, Holguin G, et al. Molecular diversity of denitrifying genes in continental margin sediments within the oxygen-deficient zone off the pacific coast of Mexico[J]. Applied and Environmental Microbiology, 2003, 69(6): 3549-3560. DOI:10.1128/AEM.69.6.3549-3560.2003 |
| [37] |
Miao Y, Liao RH, Zhang XX, et al. Metagenomic insights into salinity effect on diversity and abundance of denitrifying bacteria and genes in an expanded granular sludge bed reactor treating high-nitrate wastewater[J]. Chemical Engineering Journal, 2015, 277: 116-123. DOI:10.1016/j.cej.2015.04.125 |
| [38] |
Ma LH, Huang YM, Li XZ, et al. Changes of nitrogen forms and microbial physiological group diversity and their relations during composting of cow manure[J]. Journal of Agro-Environment Science, 2009, 28(12): 2674-2679. (in Chinese) 马丽红, 黄懿梅, 李学章, 等. 牛粪堆肥化中氮素形态与微生物生理群的动态变化和耦合关系[J]. 农业环境科学学报, 2009, 28(12): 2674-2679. DOI:10.3321/j.issn:1672-2043.2009.12.037 |