 2018, Vol. 45
2018, Vol. 45扩展功能
文章信息
- 李威, 肖熙鸥, 李可, 高晓敏, 吕玲玲
- LI Wei, XIAO Xi-Ou, LI Ke, GAO Xiao-Min, Lü Ling-Ling
- 茄子青枯病拮抗放线菌XL-6的筛选、鉴定及发酵条件优化
- Screening, identification and fermentation optimization of an antagonistic actinomycete strain XL-6 against Ralstonia solanacearum
- 微生物学通报, 2018, 45(2): 357-367
- Microbiology China, 2018, 45(2): 357-367
- DOI: 10.13344/j.microbiol.china.170191
-
文章历史
- 收稿日期: 2017-03-07
- 接受日期: 2017-05-15
- 网络首发日期(www.cnki.net): 2017-06-27

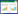




青枯病是一种由青枯菌(Ralstonia solanacearum)引起的主要作用于茄科作物的细菌性土传病害,能侵染450多种单子叶和双子叶植物[1],严重影响热带、亚热带和温带地区的农业生产。青枯病病原菌变异大、寄主范围广,至今尚无有效的防治办法[2],而化学农药仍是防治青枯病的主要措施。利用化学农药防治,不但用药量大、药效低、破坏土壤的微生态平衡、农药残留在土壤中造成污染,而且易使病原菌产生抗药性。同时,随着人们的生态意识和绿色健康环保理念的加强,使用环保、无残留的生物防治已成为共识和研究热点。
放线菌是一类潜力巨大的微生物种群,在适宜的条件下,放线菌分泌的代谢产物不但能抑制病原菌,而且能促进植物生长。目前从微生物中发现的8 000多种生物活性物质中,近2/3是由放线菌产生的[3]。抗生素作为放线菌代谢的重要活性物质已被广泛应用于医药和农业,在放线菌众多的种群中,以链霉菌产生的抗生素种类最多,占总数的50%[4]。利用放线菌次生代谢产物制备的生物农药,因其具有无毒、无污染、不易产生抗药性等特点,已成为未来农药的发展方向。在农业病害防治方面,研究人员已从土壤和植物体内筛选出不同的放线菌菌株用于对草坪草根腐病[5]、稻瘟病[6]、水稻叶枯病[7]和番茄灰霉病[8]的生物防治并获得较好的效果。同时,对拮抗放线菌代谢产物中具有抑菌作用的活性物质进行了初步的分离,兰鑫等从拮抗放线菌Hhs.015发酵液中分离并鉴定出对苹果树腐烂病菌有抑制作用的活性物质为苯乙酸[9];Roy等从链霉菌发酵液分离出对G+、G-细菌及单细胞和丝状真菌都有较强抑制作用的DBP[10]。这些研究结果为茄子青枯病的生物防治提供了新的思路和途径。
本研究从茄子发病严重地块的健康植株根系中筛选出一株对青枯菌有较强拮抗作用的放线菌菌株,对其种属关系和发酵条件进行研究,为抗生物质的分离鉴定提供实验依据,也为后期生产应用提供理论基础。
1 材料与方法 1.1 材料 1.1.1 样品来源2015年11月从广东省湛江市徐闻县西连镇英邱村茄子青枯病发病严重地块中对健康植株的根际土壤进行取样,共采集土样23份。采集土样时,取健康植株根部10-30 cm处的根际土壤并装入无菌塑料袋中,4 ℃保存。
1.1.2 培养基发酵基础培养基(g/L):葡萄糖10.0,可溶性淀粉10.0,酵母粉8.0,NaCl 0.5,K2HPO4 1.0,MgCl2 2.0,蒸馏水1 000 mL,调节pH 7.2-7.4。
1.1.3 主要试剂和仪器蛋白胨、桑塔氏培养基、溶菌酶及RNA提取试剂盒等均购自TaKaRa公司。光学显微镜,江南光学仪器厂;PCR扩增仪,TaKaRa公司。
1.2 方法 1.2.1 放线菌发酵培养基及其无菌发酵液的制备种子液:将10 µL放线菌菌悬液接种于50 mL的发酵基础培养基中,28 ℃、180 r/min振荡培养36 h制成种子液。
发酵:在发酵基础培养基中以10%的接种量接入种子液,28 ℃、180 r/min振荡培养6 d后,10 000 r/min离心10 min,经0.22 µm的滤膜过滤,得发酵滤液。
1.2.2 放线菌的分离与纯化取10 g土样放入90 mL无菌水中,150 r/min振荡30 min后依次稀释成浓度为10-3、10-4、10-5的土悬液并涂布于高氏一号培养基。28 ℃培养21 d,挑取菌落形态各不相同的菌株转接到新的高氏一号培养基上培养,经过多次单菌落纯化之后,转接到高氏一号斜面培养基上培养21 d,编号保存。
1.2.3 拮抗放线菌的筛选(1) 拮抗放线菌的初筛。采用双层琼脂法[11]对分离到的放线菌进行抗菌活性初筛。将供试放线菌培养5 d后,用打孔器(Φ=8 mm)将其切成菌块,移至高氏一号培养基中央,28 ℃培养2 d。将供试茄子青枯病菌(Ralstonia solanacearum)用无菌水配制成菌悬液(6×108 CFU/mL),用喷雾器均匀地喷洒在培养了2 d的放线菌平板上,然后倒入约15 mL温度为40 ℃左右的NA培养基制成含菌平板,每个处理重复3次,以不接种放线菌的平板作为对照,28 ℃培养24 h测量抑菌圈直径。
(2) 拮抗放线菌的复筛。选择在初筛中拮抗性好的放线菌并获得其无菌发酵液,采用琼脂扩散法[12]测定拮抗放线菌的抑菌活性。将茄子青枯菌加入NA培养基中制成浓度为4×108 CFU/mL的含菌平板,用打孔器(Φ=8 mm)在含菌平板中央打孔并取出菌块。向每个孔中注入100 µL的无菌发酵液,28 ℃培养24 h。每个处理重复3次,用十字交叉法测量抑菌圈直径大小,以注入无菌水的平板作为对照。
(3) 拮抗放线菌对其他作物土传病原菌抑制作用的测定。采用平板对峙法[13],将筛选出的拮抗放线菌培养5 d,用无菌打孔器(Φ=8 mm)打制菌饼,菌面朝下放置于PDA平板中央,28 ℃培养2 d后,再于距放线菌菌块两侧2 cm对称接种用同样方法打制的病原菌菌饼,28 ℃培养7 d。每处理重复3次。测试的病原菌为:黄瓜枯萎病原菌(Fusarium oxysporum f. sp. cucumerinum)、香蕉枯萎病原菌(Fusarium oxysporum f. sp. cubense)和茄子黄萎病原菌(Verticillium dahlia)。
1.2.4 菌株的鉴定目标菌株XL-6的鉴定工作委托中国科学院微生物研究所完成。
(1) 形态学特征:接种放线菌到高氏一号培养基中,采用插片法28 ℃培养21 d后,在光学显微镜下观察基内菌丝及气生菌丝的形状、孢子链的有无等。
(2) 培养特征及生理生化特征:参照阮继生等[14]和Shirling等[15]的方法,目标放线菌在28 ℃培养21 d,观察记录基内菌丝和气生菌丝的颜色、可溶性色素的有无等。
(3) 系统发育学特征:采用16S rRNA基因的通用引物[16]27F和1492R进行PCR扩增,测定其序列。通过BLAST进行比较,从GenBank数据库中选取相似性最高且具有代表性的菌株,用ClustalX进行序列比对,用MEGA 5.05的Neighbor-Joining法构建系统进化树,确定放线菌的分类地位[17]。
1.2.5 发酵条件的优化(1) 不同碳源、氮源对拮抗放线菌XL-6发酵液抑菌活性的影响:在培养条件不变的情况下,分别以20 g/L的葡萄糖、蔗糖、麦芽糖、果糖、甘露醇、玉米粉和甘油替代发酵基础培养基中的可溶性淀粉,分别以2 g/L的牛肉膏、蛋白胨、黄豆粉、酵母粉和尿素替代发酵基础培养基中的KNO3,按照1.2.3(3)的方法测定,重复3次,确定最佳碳源和氮源。
(2) 营养条件的正交试验:选择最佳碳源(玉米粉)和氮源(酵母粉)以及3种无机盐K2HPO4、NaCl、MgCl2共5个因素,设置4个水平,选用L16(45)正交表进行正交试验(表 1),发酵试验重复3次,抑菌活性按照1.2.3(3)的方法测定。
| 因子Factors | 水平Levels | |||
| 1 | 2 | 3 | 4 | |
| A Cornmeal (g/L) | 20.00 | 30.00 | 40.00 | 50.00 |
| B Yeast powder (g/L) | 2.00 | 3.00 | 4.00 | 5.00 |
| C K2HPO4 (g/L) | 0.25 | 0.50 | 1.00 | 2.00 |
| D MgCl2 (g/L) | 1.50 | 2.00 | 2.50 | 3.00 |
| E NaCl (g/L) | 1.00 | 1.50 | 2.00 | 2.50 |
(3) 培养条件的优化:在优化的发酵培养基基础上,在其他培养条件一致的情况下,测定不同培养条件下的发酵滤液对茄子青枯菌的抑菌效果。1)起始pH值对发酵液抑菌活性的影响:分别调节培养基的起始pH值为5.5、6.0、6.5、7.0、7.5、8.0、8.5,28 ℃、180 r/min振荡培养6 d后,获得发酵滤液,检测抑菌效果,确定最适pH值;2)瓶装量对发酵液抑菌活性的影响:在250 mL的三角瓶中,分别装入50、60、70、80、90、100、110和120 mL的培养基,28 ℃、180 r/min振荡培养6 d后,获得发酵滤液,检测抑菌效果,确定最适装瓶量;3)发酵温度对发酵液抑菌活性的影响:分别在20、25、28、30、32、37和40 ℃进行发酵培养,180 r/min振荡培养6 d后,检测发酵液抑菌效果,确定最适培养温度;4)发酵时间对发酵液抑菌活性的影响:分别培养3、4、5、6、7、8和9 d后,检测发酵液的抑菌效果,确定最适发酵时间;5)摇床转速对发酵液抑菌活性的影响:在120、140、160、180、200和220 r/min条件下,28 ℃培养6 d后检测抑菌效果,确定最适转速;6)接种量对发酵液抑菌活性的影响:将种子液以2%、4%、6%、8%、10%和12%的接种量加入到基础发酵培养基中,28 ℃、180 r/min振荡培养6 d后,检测抑菌效果,确定最适接种量。
1.2.6 数据处理利用SPSS 11.5数据软件对相关数据进行统计分析。
2 结果与分析 2.1 拮抗放线菌的分离和纯化从采集到的23份土样中共分离到59株放线菌,根据菌落形态及是否产色素,最终获得25株不同放线菌,编号为XL-1–25。25株放线菌中,大部分菌株在高氏一号培养基中不产色素,少部分菌株产浅褐和浅黄色素,1株产浅粉色素。
2.2 拮抗放线菌的初筛如图 1所示,利用平板对峙法筛选出1株对茄子青枯病原菌菌、黄瓜枯萎病原菌、香蕉枯萎病原菌和茄子黄萎病原菌均具有较强抑制作用的拮抗放线菌,编号为XL-6。

|
| 图 1 菌株XL-6对部分病原菌的抑制效果 Figure 1 Inhibition effect of strain XL-6 on some pathogen 注:A:茄子青枯病菌;B:黄瓜枯萎病菌;C:香蕉枯萎病菌;D:茄子黄萎病菌. Note: A: Ralstonia solanacearum; B: Fusarium oxysporum f. sp. cucumerinum; C: Fusarium oxysporum f. sp. cubense; D: Verticillium dahlia. |
|
|
采用琼脂扩散法测定不同浓度无菌发酵液对茄子青枯菌的抑制效果,结果如图 2所示,目标菌株XL-6的发酵液对青枯菌具有较好的抑制作用,其抑菌效果随着发酵液浓度的降低而减弱。以生产上防治青枯病的主要药物农用链霉素为对照(A),发现菌株发酵液原液(D)的抑制效果要优于农用链霉素,说明菌株XL-6具有较好的生防应用潜力。

|
| 图 2 不同浓度的菌株XL-6发酵液对青枯菌的抑制效果 Figure 2 Inhibition effect of different concentrations fermentation filtrate of strain XL-6 on Ralstonia solanacearum 注:A:农用链霉素;B:5倍发酵液;C:10倍发酵液;D:发酵原液. Note: A: Streptomycin; B: Fermentation diluted 5 times; C: Fermentation diluted 10 times; D: Fermentation filtrate. |
|
|
利用插片法培养拮抗放线菌,通过光学显微镜观察可知:在高氏一号培养基上,XL-6菌株的孢子丝直、柔曲、松敞螺旋形;孢子柱形、椭圆形(图 3)。菌株XL-6在8种供试培养基上均生长良好,其气生菌丝主要以灰色和灰白色为主,其中高氏一号培养基和甘油天冬素培养基上分别为粉灰色和豆汁黄;菌株的基内菌丝在不同培养基中呈现较为丰富的颜色,其中,在葡萄糖天冬素培养基、ISP-2培养基和桑塔氏培养基上呈软木黄色,在察氏培养基上为水牛灰色,在燕麦粉培养基上为珠母灰,在高氏一号培养基中呈现落英淡粉色;除在高氏一号培养基上生长时产落英淡粉色素外,在其他培养基上均不产色素(表 2)。

|
| 图 3 菌株XL-6的孢子链及孢子形态(4 000×) Figure 3 Morphologic characters of spore chain and spore of strain XL-6 (4 000×) |
|
|
| 培养基 Media |
气生菌丝 Aerial mycelium |
基内菌丝 Substrate mycelium |
可溶性色素 Soluble pigment |
| Czapek’s medium | Gray | Buff gray | None |
| Glucose asparagine agar | Offwhite | Light yellow | None |
| Glycerol asparagine agar | Bean yellow | Light bean yellow | None |
| Mineral salt starch agar | Lotus white | Bean yellow | None |
| ISP-2 | Offwhite | Light yellow | None |
| Oat agar | Gray | Light gray | None |
| Gause’s synthetic medium | Pink gray | Light pink | Light pink |
| Santa’s agar | Offwhite | Light yellow | None |
放线菌XL-6可使明胶液化、硝酸盐还原和牛奶胨化凝固,同时可水解淀粉(产淀粉酶)和酪氨酸(产酪氨基酸酶);对碳源的利用结果表明,菌株XL-6可利用葡萄糖、甘露糖、甘露醇、乳糖、半乳糖、麦芽糖、蔗糖、密二糖、鼠李糖、棉籽糖、海藻糖、果糖、木糖、菊糖、糖原、肌醇、甘油、苹果酸钠、琥珀酸钠及葡萄糖酸钠,不能利用山梨醇、山梨糖、松三糖、核糖及柠檬酸钠。对氮源的利用实验表明,该菌株可利用L-丙氨酸,但不能利用L-精氨酸、L-甲硫氨酸和L-赖氨酸(表 3)。
| 特征 Characteristics |
结果 Results |
| Glucose | + |
| Mannose | + |
| Mannitol | + |
| Lactose | + |
| Galactose | + |
| Sorbose | - |
| Sorbitol | - |
| Maltose | + |
| Sucrose | + |
| Melibiose | + |
| Melezitose | - |
| Rhamnose | + |
| Raffinose | + |
| Trehalose | + |
| Fructose | + |
| Xylose | + |
| Ribose | - |
| Inulin | + |
| Glycogen | + |
| Galactitol | - |
| Salicin | + |
| Amygdalin | + |
| Erythritol | - |
| Inositol | + |
| Glycerol | + |
| Sodium gluconate | + |
| Sodium malate | + |
| Sodium citrate | - |
| Sodium succinate | + |
| L-Alanine | + |
| L-Arginine | - |
| L-Methionine | - |
| L-Lysine | - |
| Nitrate reduciton | + |
| Gelatin liquefaction | + |
| Milk peptonization | + |
| Tyrosinase | + |
| Amylase | + |
| Note: +: Positive reaction; -: Negative result. | |
对菌株XL-6的16S rRNA基因序列扩增产物进行测序,得到1 404 bp序列,GenBank登录号为KY658459。选取一致性较高且已命名菌株的相关序列信息进行系统发育分析,用MEGA 5.05构建系统进化树(图 4),可知菌株XL-6与Streptomyces rochei (AB184237)聚于同一分支且序列相似性较高,相似性为99%。根据拮抗菌株XL-6的菌体形态、培养特征及生理生化特征测定结果,并结合基于16S rRNA基因的系统发育分析,将该目标菌株XL-6鉴定为娄彻氏链霉菌。

|
| 图 4 菌株XL-6基于16S rRNA基因序列的系统发育树 Figure 4 Phylogenetic tree of strain XL-6 based on 16S rRNA gene sequences 注:分支上的数字表示构建系统树时1 000次计算时形成该节点的百分比;括号内数值为GenBank登录号;标尺0.001代表 0.2%的16S rRNA基因序列的进化差异. Note: The bootstrap values (%) presented at the branches were calculated from 1 000 replications; Numbers in parentheses are GenBank accession numbers; The scale bar 0.001 represents 2 nucleotide substitutions per 1 000 nucleotides. |
|
|
由表 4可知,分别以8种不同物质作为碳源获得的发酵液对茄子青枯菌都有一定的抑制效果,其中以玉米粉作为碳源的抑菌效果最好,抑菌圈直径达13.35 mm,显著优于基础发酵培养基中以可溶性淀粉为碳源的抑菌效果(10.55 mm),而葡萄糖、果糖、蔗糖、麦芽糖、甘油及甘露醇作为碳源的抑菌效果较差,因此,优选玉米粉作为菌株XL-6发酵培养的碳源。对于氮源的筛选,除尿素外,其他6种氮源物质的发酵液对青枯菌均有一定的抑制效果。其中以酵母粉为氮源物质的抑菌效果最好,抑菌圈直径为13.25 mm,其次为蛋白胨和牛肉膏,三者的抑菌效果均显著高于基础发酵培养基中以KNO3为氮源的抑菌效果(8.36 mm),而氯化铵和黄豆粉的效果较差。因此,优选酵母粉作为发酵培养基的氮源物质。
| 碳源 Carbon source |
抑菌圈直径 Inhibition zone diameter (mm) |
| Glucose | 6.85e |
| Fructose | 7.46d |
| Sucrose | 4.25f |
| Maltose | 8.59c |
| Cornmeal | 13.35a |
| Mannose | 6.83e |
| Glycerol | 7.45d |
| Soluble starch | 10.55b |
| Beef extract | 9.50c |
| Peptone | 11.00b |
| Yeast powder | 13.25a |
| Urea | 0 |
| Soybean meal | 5.38f |
| NH4Cl | 7.05e |
| KNO3 | 8.36d |
| 注:同列数据后不同小字母表示经Duncan氏新复极差法检验,在P < 0.05水平差异显著. Note: Different lowercase letters in the same column show the significantly different at P < 0.05 level by Duncan’s new multiple range test, respectively. |
|
以玉米粉(A)、酵母粉(B)、K2HPO4 (C)、MgCl2 (D)和NaCl (E)为因素,按L16(45)设计5因素4水平的正交试验,测定菌株XL-6的发酵液对青枯菌的抑菌活性,从而确定该菌株的发酵培养基配方。
正交试验的极差分析表明,5个因素对青枯菌的抑制作用为A > D > B > E > C,最佳水平组合为A2B4C4D2E1。方差分析结果表明,因素A对青枯菌的影响显著(表 5)。因此,确定培养基配方(g/L):玉米粉30.0、酵母粉5.0、K2HPO4 2.0、MgCl2 2.0和NaCl 1.0。利用最佳水平组合A2B4C4D2E1的发酵培养基发酵,获得的无菌发酵液对青枯菌的抑菌圈直径达19.33 cm,比实际优化后的抑菌效果(抑菌圈直径17.18 cm)提高了12.51%。
| 序号 Number |
A | B | C | D | E | 抑菌圈直径 Inhibition zone diameter (mm) |
| 1 | 1 | 1 | 1 | 1 | 1 | 9.21 |
| 2 | 1 | 2 | 2 | 2 | 2 | 10.63 |
| 3 | 1 | 3 | 3 | 3 | 3 | 8.23 |
| 4 | 1 | 4 | 4 | 4 | 4 | 8.93 |
| 5 | 2 | 1 | 2 | 3 | 4 | 11.94 |
| 6 | 2 | 2 | 1 | 4 | 3 | 13.13 |
| 7 | 2 | 3 | 4 | 1 | 2 | 15.05 |
| 8 | 2 | 4 | 3 | 2 | 1 | 17.18 |
| 9 | 3 | 1 | 3 | 4 | 2 | 7.91 |
| 10 | 3 | 2 | 4 | 3 | 1 | 12.04 |
| 11 | 3 | 3 | 1 | 2 | 4 | 13.76 |
| 12 | 3 | 4 | 2 | 1 | 3 | 12.29 |
| 13 | 4 | 1 | 4 | 2 | 3 | 10.19 |
| 14 | 4 | 2 | 3 | 1 | 4 | 8.10 |
| 15 | 4 | 3 | 2 | 4 | 1 | 8.44 |
| 16 | 4 | 4 | 1 | 3 | 2 | 8.14 |
| X1 | 9.25 | 9.81 | 11.06 | 11.16 | 1.72 | |
| X2 | 14.33 | 10.98 | 10.83 | 12.94 | 0.43 | |
| X3 | 11.50 | 11.37 | 10.36 | 10.09 | 0.96 | |
| X4 | 8.72 | 11.64 | 11.55 | 9.60 | 0.68 | |
| R | 5.61 | 1.82 | 1.20 | 3.34 | 1.28 |
在确定了发酵培养基的基础上,对目标菌株的发酵条件进行了优化。由图 5可知,适宜目标菌株产抑菌活性物质的发酵培养条件为:初始pH为7.0,培养基装瓶量为70 mL/250 mL,摇床转速为180 r/min,接种量为6%,在28 ℃条件下培养6 d。

|
| 图 5 不同培养条件对菌株XL-6发酵液抑菌活性的影响 Figure 5 Effect of different condition on inhibition activity of submerged cultured XL-6 注:A:不同pH;B:不同装液量;C:不同温度;D:不同时间;E:不同转速;F:不同接种量.不同小字母表示经Duncan氏新复极差法检验,在P < 0.05水平差异显著. Note: A: Different pH; B: Different bottle filling quantity; C: Different temperature; D: Different time; E: Different rotate speed; F: Different inoculation amount. Different lowercase letters show the significantly different at P < 0.05 level by Duncan's new multiple range test, respectively. |
|
|
放线菌作为生物防治中一类重要的微生物类群,可分泌抗生素、生物碱、酶制剂等直接抑制病原菌的生长,也可与其他病原菌之间形成营养和空间的竞争关系而发挥拮抗抑菌作用,或通过分泌植物促生物质以提高植物抗性,从而达到防治效果。本研究从发病严重地块的健康茄子植株根际分离出一株对茄子青枯菌有较强抑制作用的放线菌菌株,命名为XL-6。采用传统分类法和16S rRNA基因序列相结合初步鉴定菌株XL-6为娄彻氏链霉菌,与数据库中已登录的菌株ZZ-9 (Streptomyces rochei KT986228)[18]、Fro2 (Streptomyces rochei KC522302)[19]和D164 (Streptomyces rochei JQ901942)[20]等其他娄彻氏链霉菌相比,在生理生化特征上有较小的差异。
娄彻氏链霉菌广泛存在于自然界中,其用来防治植物病害已有相关报道。相关研究已在黄瓜枯萎病[21]、棉花黄萎病[22]、稻瘟病[23]、金黄色葡萄球菌[24]上开展并取得了初步成效。此外,娄彻氏链霉菌不仅能够产生多种具有抗菌活性的次级代谢产物,而且其自身还具有很高的抗植物病原菌、杀虫及除草活性,并在植物病毒和植物病原真菌的生物防治中都有较高的潜在应用价值[25]。通过对拮抗放线菌XL-6广谱抗性的研究发现,该菌株不但对青枯菌有较强的抑制作用,而且对黄瓜枯萎病菌、香蕉枯萎病菌1号生理小种、香蕉枯萎病菌4号生理小种、茄子黄萎病菌等土传病原菌均有较好的抑制作用,说明该菌株具有广谱的抗菌特性,在以后的生产应用中具有较大的潜力。但目前的研究表明田间环境和植物微生态环境是影响生防菌在田间发挥抑病作用的重要影响因素[26]。因此,为了更好地将生防菌株XL-6用于实际生产,后续研究需对发酵液中抑菌活性物质进行分离鉴定,对抑菌机理和田间防效等进行深入的探索。
抗菌活性物质通常作为生防菌发挥生物防治作用的物质基础,其产量的高低影响着生防菌株的实际应用效果[27]。发酵是获得大量微生物活性代谢产物的基础,微生物代谢产物的类型和产量与其培养条件,如培养基中的碳源、氮源、pH等密切相关[28],因此探索出适宜的培养条件尤为关键,直接关系到目标生防菌株的实际应用价值。本研究采用单因素和正交试验进行菌株XL-6发酵条件的优化,初步获得了适宜目标菌株产抑菌活性物质的发酵营养基配方和培养条件,优化后发酵液的抑菌活性显著高于基础发酵培养基,但在本研究中未考虑交互作用和发酵成本,所以应在此优化培养基的基础上,兼顾成本和交互作用的影响,得到更好的发酵条件,以便于推广应用。
微生物发酵是个动态的生物学过程,培养基中各养分含量、pH、容氧量和产物量不断变化,不同因素的变化对发酵产物的种类和产量均有较大的影响,而在本研究中仅考虑了发酵前的培养基状态和培养条件,未对发酵过程各因素进行动态监控。在后续的研究中,应采用生物化学、分子生物学等方法对目标菌株的代谢动态过程进行深入研究,探讨培养基的各养分含量与目标活性物质产量及培养条件之间的关系,进而提高目标活性物质的产率。
致谢: 本研究中目标拮抗菌XL-6的生理生化鉴定及16S rRNA基因序列分析均由中国科学院微生物研究所完成,在此表示衷心的感谢。| [1] |
Wicker E, Grassart L, Coranson-Beaudu R, et al. Ralstonia solanacearum strains from Martinique (French West Indies) exhibiting a new pathogenic potential[J]. Applied and Environmental Microbiology, 2007, 73(21): 6790-6801. DOI:10.1128/AEM.00841-07 |
| [2] |
Yin XG, Wang XJ, Zhang Y, et al. Research progress on tomato bacterial wilt and resistance breeding in China[J]. Journal of Yunnan Agricultural University, 2005, 20(2): 163-167. 尹贤贵, 王小佳, 张赟, 等. 我国番茄青枯病及抗病育种研究进展[J]. 云南农业大学学报, 2005, 20(2): 163-167. |
| [3] |
Jiang Y, Huang LL, Chen CQ, et al. Screen, identification and optimized fermentation condition of an actinomycete strain against pathogenic fungus Fulvia fulva[J]. Acta Microbiologica Sinica, 2007, 47(4): 622-627. 姜云, 黄丽丽, 陈长卿, 等. 一株拮抗番茄叶霉病菌的放线菌筛选、鉴定及发酵条件研究[J]. 微生物学报, 2007, 47(4): 622-627. |
| [4] |
McKenzie NL, Thaker M, Koteva K, et al. Induction of antimicrobial activities in heterologous streptomycetes using alleles of the Streptomyces coelicolor gene absA1[J]. The Journal of Antibiotics, 2010, 63(4): 177-182. DOI:10.1038/ja.2010.13 |
| [5] |
Wang Q, Duan BH, Yang R, et al. Screening and identification of chitinolytic actinomycetes and the inhibitory activity on turfgrass root rot disease fungi[J]. Acta Agrestia Sinica, 2013, 21(5): 998-1004. 王歧, 段碧华, 杨瑞, 等. 产几丁质酶放线菌的筛选、鉴定及其对草坪草根腐病真菌的抑制作用[J]. 草地学报, 2013, 21(5): 998-1004. DOI:10.11733/j.issn.1007-0435.2013.05.025 |
| [6] |
Wang ZZ, Xu T, Yuan SS, et al. Identification of an endophytic actinomyce OsiRt-1 isolated from rice and its effect against rice blast disease[J]. Microbiology China, 2016, 43(5): 1009-1018. 王真真, 徐婷, 袁姗姗, 等. 水稻内生放线菌OsiRt-1的分离鉴定及对稻瘟病的防治作用[J]. 微生物学通报, 2016, 43(5): 1009-1018. |
| [7] |
van Hop D, Hoa PTP, Quang ND, et al. Biological control of Xanthomonas oryzae pv. oryzae causing rice bacterial blight disease by Streptomyces toxytricini VN08-A-12, isolated from soil and leaf-litter samples in Vietnam[J]. Biocontrol Science, 2014, 19(3): 103-111. DOI:10.4265/bio.19.103 |
| [8] |
Kobayashi YO, Kobayashi A, Maeda M, et al. Biological control of potato scab and antibiosis by antagonistic Streptomyces sp. WoRs-501[J]. Journal of General Plant Pathology, 2015, 81(6): 439-448. DOI:10.1007/s10327-015-0614-y |
| [9] |
Lan X, Yan X, Zhen W, et al. Studies on the anti-microbial active substances from the fermentation broth of cucumber endophytic actinomyce Hhs.015[J]. Journal of Agricultural University of Hebei, 2015, 38(6): 73-78. 兰鑫, 颜霞, 甄伟, 等. 黄瓜内生放线菌Hhs.015发酵液抑菌活性成分的研究[J]. 河北农业大学学报, 2015, 38(6): 73-78. |
| [10] |
Roy RN, Laskar S, Sen SK. Dibutyl phthalate, the bioactive compound produced by Streptomyces albidoflavus 321.2[J]. Microbiological Research, 2006, 161(2): 121-126. DOI:10.1016/j.micres.2005.06.007 |
| [11] |
Stonier T. Agrobacterium tumefaciens Conn Ⅱ. Production of an antibiotic substance[J]. Journal of Bacteriology, 1960, 79(6): 889-898. |
| [12] |
Zhou DQ. Microbiology Tutorial[M]. Beijing: Higher Education Press, 2002, 2nd ed: 121-124. 周德庆. 微生物学教程[M]. 北京: 高等教育出版社, 2002, 2版: 121-124. |
| [13] |
Sijam K, Dikin A. Biochemical and physiological characterization of Burkholderia cepacia as biological control agent[J]. International Journal of Agriculture & Biology, 2005, 7(3): 385-388. |
| [14] |
Ruan JS, Liu ZH, Liang LN. Actinomycetes Research and Application[M]. Beijing: Science Press, 1990, 77-79. 阮继生, 刘志恒, 梁丽糯. 放线菌研究及应用[M]. 北京: 科学出版社, 1990, 77-79. |
| [15] |
Shirling EB, Gottlieb D. Methods for characterization of Streptomyces species[J]. International Journal of Systematic and Bacteriology, 1966, 16(3): 313-340. DOI:10.1099/00207713-16-3-313 |
| [16] |
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT[J]. Nucleic Acids Symposium Series, 1999, 41: 95-98. |
| [17] |
Wang J, Huang Y, Yao J, et al. Identification and control effects of two antagonistic actinomycetes against clubroot[J]. Scientia Agricultura Sinica, 2011, 44(13): 2692-2700. 王靖, 黄云, 姚佳, 等. 两株根肿病生防放线菌的鉴定及其防病效果[J]. 中国农业科学, 2011, 44(13): 2692-2700. DOI:10.3864/j.issn.0578-1752.2011.13.007 |
| [18] |
Xue YY, Fan WZ, Zhang SW, et al. Screening, identification and biocontrol effect of antagonistic actinomycetes against the pathogen of Cytospora sp. for apple tree[J]. Chinese Journal of Applied Ecology, 2016, 27(10): 3379-3386. 薛应钰, 范万泽, 张树武, 等. 苹果树腐烂病菌拮抗放线菌的筛选、鉴定及防效[J]. 应用生态学报, 2016, 27(10): 3379-3386. |
| [19] |
Zhang ZB, Ao W, Xiong YY, et al. Identification of antagonistic endophytic actinomycete FRo2 and isolation of its antimicrobial composition[J]. Microbiology China, 2014, 41(8): 1574-1581. 张志斌, 敖武, 熊瑶瑶, 等. 内生拮抗放线菌FRo2的鉴定及抑菌活性物质的分离[J]. 微生物学通报, 2014, 41(8): 1574-1581. |
| [20] |
Xin YJ, Ai JN, Liu YN, et al. Isolation, identification and bioactivity screening of marine actinobacteria[J]. Current Biotechnology, 2012, 2(5): 354-358. 信艳娟, 艾江宁, 刘亚男, 等. 一株海洋放线菌的分离鉴定及活性研究[J]. 生物技术进展, 2012, 2(5): 354-358. |
| [21] |
Liu Q, Xu J, Liu HE, et al. Isolation and antagonistic activity of endophytic actinomycete SR-1102 to Fusarium oxysporum f. sp. cucumbrium of cucumber[J]. Journal of Yangzhou University (Agricultural and Life Science Edition), 2015, 36(2): 83-88. 刘琴, 徐健, 刘怀阿, 等. 黄瓜内生放线菌SR-1102分离及对枯萎病菌拮抗活性[J]. 扬州大学学报:农业与生命科学版, 2015, 36(2): 83-88. |
| [22] |
Liu CB, Wan CX, Jia XY, et al. Screening of antagonistic actionomycetes against verticillium dahliae from saline environments in Xinjiang as well as the antagonistic activities and identification of strain TRM42561[J]. Chinese Journal of Biological Control, 2014, 30(3): 408-413. 柳成宾, 万传星, 贾晓宇, 等. 新疆盐环境棉花黄萎菌拮抗放线菌的筛选及菌株TRM42561的防效测定和鉴定[J]. 中国生物防治学报, 2014, 30(3): 408-413. |
| [23] |
Zhang L, Ji MS, Yu ZG. Antifungal activities and stability of metabolites from Streptomyces rochei YL-2 against Pyricularia oryzae[J]. Journal of Shenyang Agricultural University, 2014, 45(2): 143-146. 张丽, 纪明山, 于志国. 娄彻氏链霉菌YL-2代谢产物对稻瘟病菌的抑制活性及其稳定性[J]. 沈阳农业大学学报, 2014, 45(2): 143-146. |
| [24] |
Xiong YY, Ao W, Li Y, et al. The identification of three antagonistic endophytic actinomycete and their antimicrobial activities[J]. Journal of Jiangxi Normal University (Natural Sciences Edition), 2015, 39(2): 171-176. 熊瑶瑶, 敖武, 李怡, 等. 3株拮抗内生放线菌的鉴定及其抑菌活性初步研究[J]. 江西师范大学学报:自然科学版, 2015, 39(2): 171-176. |
| [25] |
Li Z, Zhao SY. Borrelidin production by Streptomycete rochei ATCC10739: optimization of fermentation, separation and purification conditions[J]. Microbiology China, 2011, 38(7): 1051-1055. 李周, 赵圣印. 娄彻氏链霉菌ATCC10739产抗生素Borrelidin发酵条件优化及其分离纯化[J]. 微生物学通报, 2011, 38(7): 1051-1055. |
| [26] |
Pan ZB, Li QL, Mo JY, et al. Screening, identification and control efficacy of antagonistic bacteria against Colletotrichum gloeosporioides on mango[J]. Acta Phytophylacica Sinica, 2013, 40(6): 517-522. 潘朝勃, 李其利, 莫贱友, 等. 杧果炭疽菌拮抗细菌的筛选鉴定及其防病效果[J]. 植物保护学报, 2013, 40(6): 517-522. |
| [27] |
Cao QQ, Zhou DB, Zheng L, et al. Screening, identification and cultivation conditions of microbes antagonistic to rice sheath blight fungus Rhizoctonia solani[J]. Chinese Journal of Biological Control, 2013, 29(2): 270-276. 曹琦琦, 周登博, 郑丽, 等. 水稻纹枯病菌拮抗菌的筛选、鉴定及培养条件探索[J]. 中国生物防治学报, 2013, 29(2): 270-276. |
| [28] |
Lin H, Pu XM, Zhang RL, et al. Screening of the antifungal strains of actinomycetes and the study on one of the active strains[J]. Journal of Jinan University (Natural Science Edition), 2012, 33(1): 76-80. 林虹, 蒲小明, 张荣柳, 等. 放线菌抗真菌活性筛选及其中一菌株的初步研究[J]. 暨南大学学报:自然科学版, 2012, 33(1): 76-80. |