 2015, Vol. 42
2015, Vol. 42扩展功能
文章信息
- 周叶鸣, 邹晓, 瞿娇娇, 梁宗琦
- ZHOU Ye-Ming, ZOU Xiao, QU Jiao-Jiao, LIANG Zong-Qi
- 中国虫草一新记录种
- A new record of the genus Ophiocordyceps from China
- 微生物学通报, 2015, 42(9): 1710-1716
- Microbiology China, 2015, 42(9): 1710-1716
- 10.13344/j.microbiol.china.150424
-
文章历史
- 收稿日期: 2015-05-28
- 接受日期: 2015-06-29
- 优先数字出版日期(www.cnki.net): 2015-07-14

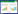




被毛孢属HirsutellaPat.是以嗜虫被毛孢Hirsutellaentomophila Pat.作为模式种而建立。该属主要特征为:孢梗束一般柱状、丝状,单生或分枝,有或无;菌丝通常无色、淡色或深褐色;产孢细胞散生于菌丝体上或者侧生于孢梗束外围菌丝上,表面光滑或粗糙,颈部简单或再育成多个小颈,颈部一般直,部分种可呈螺旋状或波浪状;分生孢子单细胞,少数多细胞,一般为柠檬形、椭圆形、梭形、橘瓣形等,经常埋于无色或有色的黏液层中;少数种还可形成二次分生孢子[1]。迄今,Index Fungorum (http://www.indexfungorum.org)收录的被毛孢共有109个分类单元。被毛孢属有4个种与本文描述的新记录种接近,并具有两型产孢细胞且锥形产孢细胞长度大于40 μm,分别是H. stilbelliformis var. stilbelliformis H. C. Evans & Samson、H. stilbelliformis var. dolichoderi H. C. Evans & Samson[2]、H. sporodochialis H. C. Evans & Samson[3]、H. Zhangjiajiensis Z. Q. Liang[4]。
被毛孢属的寄主范围很广,包括蜘蛛、螨类、昆虫等节肢动物和线虫,具有潜在的生物防治价值[5, 6, 7]。20世纪80年代早期发现H. rhossiliensis Minter & B. L. Brady对许多植物的寄生线虫如茎线虫、根结线虫等有较强的致死效果[8, 9];H. citriformis Speare对果园中的柑橘木虱有很好的防治效果[10]。被毛孢属的有性阶段为线形虫草属Ophiocordyceps(Petch) Sung et al[11],虫草是亚洲许多国家的传统药物,因此被毛孢属真菌中的活性物质也受到医药开发研究者的关注。研究表明一些被毛孢能产生各种对人类有价值的活性化合物,这些化合物具有抗肿瘤和抗疟疾的功效[12, 13]。
目前,已报道的无性型是被毛孢的线形虫草,共有27个种[11]。巨针线形虫草Ophiocordyceps macroacicularis S. Ban, T. Sakane & Nakagiri是最近被报道的一个具有被毛孢无性型的线形虫草属新种[14]。其特征为:子座丛生,圆柱形,无分枝(97.2−166.1) mm×(1.3−2.4) mm;子囊壳垂直表生,卵形,淡褐色到棕色,(410−760) μm×(260−420) μm;子囊透明,圆柱形,长235−310 μm,子囊帽显著增厚,直径5.0−7.0 μm;子囊孢子针形的,具隔,透明,(200−300) μm×(2.3−3.0) μm。同时,文中明确指出巨针线形虫草的无性阶段是被毛孢并提供了图片,但没有对无性阶段进行详细描述,也没有与其它已发表被毛孢种类进行比较。本文从形态分类鉴定的角度对其无性阶段进行了鉴定和描述,讨论了该种与被毛孢属中其它相关种的区别,为该虫草的正确分类提供了无性阶段的鉴定证据。
1 材料与方法 1.1 材料 1.1.1 标本:标本采自贵州省贵阳市桐木岭(26°23'00.3"N,106°40'33.0"E),寄主为蝠蛾科幼虫。标本编号:Dxhir140901;采集人:周叶鸣、瞿娇娇;采集时间:2014年9月。菌株编号:Gzuifr-hir140901,现保存于贵州大学真菌资源研究所菌种保藏中心(GZAC)。 1.1.2 培养基与染色液:PDA培养基:马铃薯200 g切块,用水煮沸30 min,过滤取汁,加葡萄糖20 g,水补足1 000 mL,pH自然,1×105 Pa灭菌30 min,凝固前加入链霉素和青霉素,倒平板备用。棉蓝染色液:苯胺蓝0.025 g,乳酸10 g,石炭酸10 g,甘油20 g,蒸馏水10 mL。配制后用双层搽镜纸过滤后备用。 1.1.3 分子系统学分析所用序列:将菌株Gzuifr-hir140901测序得到的ITS1-5.8S-ITS2核酸序列经NCBI的BLAST程序进行比对,下载相似度大于93%的核酸序列以及被毛孢属和线形虫草属的部分真菌序列。表 1为本研究用于分子系统学分析的序列。| 种类 species | Genbank登录号 GenBank accession No. | 种类 species | GenBank登录号 GenBank accession No. |
| Metacordyceps liangshanensis | KJ021169 | H. minnesotensis | DQ078757 |
| M. liangshanensis | KJ021172 | H. nodulosa | EF194146 |
| Cordyceps emeiensis | AJ309347 | H. proturicola | AB378557 |
| C. sinensis | AY608925 | H. rhossiliensis | AY745253 |
| C. sinensis | AY608925 | H. rostrata | EF194150 |
| Ophiocordyceps robertsii | KC167174 | H. satumaesis | KJ913066 |
| O. macroacicularis | AB968400 | H. sinensis | AJ309355 |
| O. sinensis | EU570957 | H. thompsonii | EF194153 |
| O. sinensis | JQ013106 | H. tunicata | JN247824 |
| O. stylophora | JN943352 | H. uncinata | AY365468 |
| H. aphidis | EF194156 | H. vermicola | DQ345592 |
| H. citriformis | JF894156 | H. versicolor | DQ345594 |
| H. guyana | DQ345595 | Beauveria bassiana | AF322932 |
| H. huangshanensis | EF689043 | B. bassiana | DQ364698 |
| H. liboensis | FJ957892 | Gzuifr-hir140901 | KR005829 |
巨针线形虫草的无性阶段 图 1

|
| 图 1 巨针线形虫草无性阶段产孢结构和菌落特征(Gzuifr-hir140901) Figure 1 Conidiogenous structures and colonies of the anamorph of Ophiocordyceps macroacicularis (Gzuifr- hir140901) 注:A:标本(Dxhir140901);B−E:在PDA上培养30 d后形成的菌落;F,H:产孢结构;G:分生孢子.标尺:B−E:10 mm;F−H:10 μm.Note:A:The specimen (Dxhir140901);B−E:Clonies on PDA media for 30 days;F,H:The phialides structure on hyphae;G:Conidia.B−E:Bar=10 mm;F−H:Bar=10 μm. |
The anamorph of Ophiocordyceps macroacicularis Ban, T. Sakane & Nakagiri Figure 1
The fungus hardly grow above 30 °C while growing slowly under 16 °C on the PDA agar, and colonies diam up to 25−30 mm after 30 d. The colonies margin be white and the middle light-brown part with cashmere. Mycelium is hyaline, smooth, septate, wide 3.6−4.5 μm. Two bottles of terrier type were formed, A-phialides, column, (1.8−6.3) μm×1.8 μm. B-phialides, columnar base, awl-shap and tapers, width of base 3−3.8 μm, length 21−63 μm, width of neck 1.8−2.0 μm; Phialides formed directly from the mycelium end, occasional polyphialidic. Conidia orange segments or oval, (8.1−10.8) μm×(2.7−5.4) μm, enveloped in a hyaline mucus, thickness 1.8−2.7 μm.
Teleomorph: Ophiocordyceps macroacicularis Ban, T. Sakane & Nakagiri.
Host: Larva of a species of Hepialidae, Lepidoptera.
Specimen examined: Dxhir140901, 2014, Y. M. Zhou & J. J. Qu collected from the Tongmuling of Guiyang city, Guizhou province and Gzuifr-hir140901, strains were isolated by Y. M. Zhou conserved in the Insititute of fungal resource, Guizhou University.
菌落:在PDA上,30 °C以上不生长,16 °C培养30 d菌落直径25−30 mm。毡状,边缘白色,中间灰褐色有凸起,凸起处有绒毛,背面浅褐色;菌丝:透明、光滑、具隔,宽3.6−4.5 μm;产孢细胞:与菌丝无明显区分;形成两种类型产孢细胞,A型:柱状,(1.8−6.3) μm×1.8 μm。B型:锥形,基部柱状宽3−3.8 μm,向上逐渐变细,长21−63 μm,颈部宽1.8−2.0 μm。菌丝末端可形成产孢细胞,有再育现象;分生孢子:橘瓣状或卵型,(8.1−10.8) μm×(2.7−5.4) μm,孢子外具粘液,厚度为1.8−2.7 μm。
有性阶段:Ophiocordyceps macroacicularis S. Ban,T. Sakane & Nakagiri。
寄主:鳞翅目,蝠蛾科幼虫。
标本:标本Dxhir140901及其分离株Gzuifr- hir140901均保存于贵州大学真菌资源研究所菌种保藏中心(GZAC)。
2.2 分子系统学与拆分网络分析结果系统发育分析(图 2)发现,菌株Gzuifr- hir140901与Ophiocordyceps macroacicularis聚在同一分支,支持率为98%,且与其他的被毛孢相比又独立分支。用Splits Tree 4构建的拆分网络图(图 3)展示了与系统发育树相似的结果。在网络中连接Gzuifr-hir140901的拆分图像,表明了它与Ophiocordyceps macroacicularis基本相同且与相关群存在明显差别。折分网络分析支持了系统发育树(图 2)展示的结果。系统发育树(图 2) A分支中的线形虫草的无性型皆为被毛孢,且基本都寄生于鳞翅目幼虫,寄生线虫的H. minnesotensis Minter & B. L. Brady和分离自植物的H. uncinata Seifert & H. Boulay未见有关它们有性阶段的报道;B分支的被毛孢属种类具有一个共同的形态学特征,它们的产孢细胞颈部具有螺旋。

|
| 图 2 菌株Gzuifr-hir140901及其相关种类基于ITS1-5.8S-ITS2的系统发育树 Figure 2 Phylogenetic tree based on rDNA ITS1-5.8S-ITS2 sequences of Gzuifr-hir140901 related species |

|
| 图 3 基于图2中A分支序列构建的无根Nieghor-Net网络图 Figure 3 Reconstructure of the Nieghor-Net network from taxa of A in Figure 2 |
Evans & Samson (1982, 1984)报道了一些具有两型产孢细胞的种,A型在菌丝的侧面生长,菌丝与产孢细胞之间有一个压实的层;B型(锥形)多长在菌丝的末端[2, 3]。拥有锥形产孢细胞且长度最大的、大于40 μm的被毛孢(表 2)有Hirsutella aphidis petch[13],H. besseyi E. E. Fisher[17],H. darwinii Evans & Samson[2],H. guignardii (Maheu) Samson et al[3],H. sinensis Liu et al[18],H. stilbelliformis var. dolichoderi H. C. Evans & Samson, H. stilbelliformis var. stilbelliformis H. C. Evans & Samson[2],H. sporodochialis H. C. Evans & Samson[3],H. zhangjiajiensis Z. Q. Liang[4],H. danubiensis Bałazy et al, H. vandergeestii Bałazy et al[19]。同时具有两型瓶梗的种为H. stilbelliformis var. stilbelliformis H. C. Evans & Samson,H. stilbelliformis var. dolichoderi H. C. Evans & Samson,H. sporodochialis H. C. Evans & Samson,H. zhangjiajiensis Z. Q. Liang。H. stilbelliformis var. stilbelliformis和H. stilbelliformis var. dolichoderi 寄生膜翅目,H. zhangjiajiensis寄生蛹。目前都没有关于短小柱状瓶梗的报道。
| 种类 species | 产孢细胞 conidiogenous cell | 孢子 Conidia (μm) | 参考文献 References | ||
| 形状 Shape | 最大长度 Maxlength (μm) | 类型 Type | |||
| Hirsutella aphidis petch | 锥形 | >40 | B | 船形9×(1.5-2.5) | [13] |
| H. besseyi Fisher | 锥形或圆柱形 | >40 | B | 椭圆形或柠檬形 (4.1-8.3)×(2.5-5.8) | [17] |
| H. danubiensis Balazy et al | 锥形 | >60 | B | 橘瓣状(4.5-6.6)×(1.9-2.7) | [19] |
| H. darwinii H. C. Evans & Samson | 锥形 | >40 | B | 梭形(4.5-11.5)×(1.5-2) | [3] |
| H. guignardii (Maheu) Samson et al | 锥形 | >40 | B | 椭圆形或梭形 (7-13)×(4-6) | [2] |
| H. sinensis Liu et al | 锥形 | >40 | B | 肾脏形或椭圆形 (5.4-1.4)×(3.2-5.4) | [18] |
| H. sporodochialis H. C. Evans & Samson | 基部膨大;瓶形或锥形 | >80 | A&B | 梭形(10-27)×(3.5-4) | [2] |
| H. stilbelliformis var. stilbelliformis H. C. Evans& Samson | 基部膨大;椭圆形有刺或锥形 | >10 >100 | A&B | 棍棒状(7-9)×(1.5-2.5) 卵形的(8-12)×(4-5) | [3] |
| H. stilbelliformis var. dolichoderi H. C. Evans & Samson | 基部膨大;椭圆形有刺或锥形 | >40 >100 | A&B | 圆柱形到卵形 (6.5-9.5)×(3.5-4.5) | [3] |
| H. vandergeestii Balazy et al | 近似锥形, 底部较厚 | >40 | B | 橘瓣状(4-) (4.2-5.5)×(2.1-2.5)(-3.0) | [19] |
| H. zhangjiajiensis Z. Q. Liang | 锥形或基部膨大;椭圆形 | >40 | A&B | 梭形或橘瓣状 (4.5-10)×(1.5-2.5) | [4] |
| Ophiocordyceps macroacicularis S. Ban, T. Sakane & Nakagiri | 近似锥形或 基部膨大 | >40 | - | 卵形到柠檬形 平均(8.1×4.9) (通过图片比例尺测量) | [14] |
| Gzuifr-hir140901 | 锥形;柱状 | >40 | A&B | 橘瓣状或卵形 (8.1-10.8)×(2.7-5.4) | 本文 |
系统发育树被广泛应用在分析、解释各级分类阶元的系统学关系,但其不能展示物种在进化过程中,杂交重组、基因水平转移和基因重复缺失等进化事件。进化网络分析可以较好地解决这个问题[20],它能提供一个传统进化树不能展示的、被平行事件掩盖的系统发育关系图像[21, 22, 23]。根据分子系统学(图 2)和系统进化(图 3)分析结果,菌株Gzuifr-hir14090可能为巨针线形虫草O.macroacicularis的无性阶段得到了很好的支持。经过与巨针线形虫草文献[14]中的无性阶段形态比较(表 2),二者基本相同。
综上所述,标本Dxhir140901及其分离菌株Gzuifr-hir140901确定为巨针线形虫草O. macroacicularis S. Ban, T. Sakane & Nakagiri的无性阶段,为中国的新记录种。
| [1] | Liang ZQ. Studies on classification of the genus Hirsutella Pat. Ⅰ. Advances and the characteristics of taxonomy[J]. Journal of Guizhou Agriculture College, 1990, 9(1): 58-68 (in Chinese) 梁宗琦. 被毛孢属(Hirsutella Pet.)的分类研究Ⅰ. 进展及分 类特征[J]. 贵州农学院学报, 1990, 9(1): 58-68 |
| [2] | Evans HC, Samson RA. Cordyceps species and their anamorphs pathogenic on ants (Formicidae) in tropical forest ecosystems Ⅱ. The Camponotus (Formicinae) complex[J]. Transactions of the British Mycological Society, 1984, 82(1): 127-150 |
| [3] | Evans HC, Samson RA. Cordyceps species and their anamorphs pathogenic on ants (Formicidae) in tropical forest ecosystems I. The Cephalotes (Myrmicinae) complex[J]. Transactions of the British Mycological Society, 1982, 79(3): 431-453 |
| [4] | Liang ZQ, Han YF, Liu AY, et al. Some entomogenous fungi from Wuyishan and Zhangjiajie Nature Reserves Three new species of the genus Hirsutella[J]. Mycotaxon, 2005, 94(5): 349-355 |
| [5] | Zou X, Liu AY, Liang ZQ, et al. Hirsutella liboensis, a new entomopathogenic species affecting Cossidae (Lepidoptera) in China[J]. Mycotaxon, 2010, 111(1): 39-44 |
| [6] | Kurihara Y, Shirouzu T, Tokumasu S, et al. Hirsutella proturicola sp. nov. isolated from a proturan, Baculentulus densus (Protura, Hexapoda)[J]. Mycoscience, 2009, 50(1): 56-62 |
| [7] | Fang HM, Tan SM. A new species of Hirsutella leizhouensis Fang & Tan[J]. Mycosystema, 1992, 11(1): 28-31 (in Chinese) 方焕谋, 谭树明. 多毛孢属一个新种—雷州多毛孢[J]. 真菌 学报, 1992, 11(1): 28-31 |
| [8] | Jaffee BA, Zehr EI. Parasitism of the nematode Criconemella xenoplax by the fungus Hirsutella rhossiliensis[J]. Phytopathology, 1982, 72(10): 1378-1381 |
| [9] | Cayrol JC, Frankowski JP. Influence of the number of parasitizing conidia of Hirsutella rhossiliensis on the mortality of Ditylenchus dipsaci[J]. Revue de Nématologie, 1986, 9(4): 411-412 |
| [10] | Hall DG, Hentz MG, Meyer JM, et al. Observations on the entomopathogenic fungus Hirsutella citriformis attacking adult Diaphorina citri (Hemiptera: Psyllidae) in a managed citrus grove[J]. BioControl, 2012, 57(5): 663-675 |
| [11] | Sung GH, Hywel-Jones NL, Sung JM, et al. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi[J]. Studies in Mycology, 2007, 57: 5-59 |
| [12] | Isaka M, Hywel-Jones NL, Somrithipol S, et al. Antituberculosis compounds, Hirsutellones A, B, and C: U. S. Patent 7, 414, 069[P]. 2008-8-19 |
| [13] | Thongtan J, Saenboonrueng J, Rachtawee P, et al. An antimalarial tetrapeptide from the entomopathogenic fungus Hirsutella sp. BCC1528[J]. Journal of Natural Products, 2006, 69(4): 713-714 |
| [14] | Ban S, Sakane T, Nakagiri A. Three new species of Ophiocordyceps and overview of anamorph types in the genus and the family Ophiocordyceptaceae[J]. Mycological Progress, 2015, 14(1): 1-12 |
| [15] | Tigano-Milani MS, Samson RA, Martins I, et al. DNA markers for differentiating isolates of Paecilomyces lilacinus[J]. Microbiology, 1995, 141(1): 239-245 |
| [16] | Kumar S, Tamura K, Nei M. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment[J]. Briefings in Bioinformatics, 2004, 5(2): 150-163 |
| [17] | Minter DW, Brady BL, Hall RA. Five hyphomycetes isolated from Eriophyid mites[J]. Transactions of the British Mycological Society, 1983, 81(3): 455-471 |
| [18] | Liu XJ, Guo YL, Yu YX, et al. Isolation and identification of the anamorphic state of Cordyceps sinensis (Berk.) Sacc[J]. Mycosystema, 1989, 8(1): 35-40 (in Chinese) 刘锡琏, 郭英兰, 俞永信, 等. 冬虫夏草菌无性阶段的分离 和鉴定[J]. 真菌学报, 1989, 8(1): 35-40 |
| [19] | Bałazy S, Miętkiewski R, Tkaczuk C, et al. Diversity of acaropathogenic fungi in Poland and other European countries[J]. Experimental and Applied Acarology, 2008, 46(1/4): 53-70 |
| [20] | Morrison DA. Phylogenetic networks: a review of methods to display evolutionary history[J]. Annual Research & Review in Biology, 2014, 4(10): 1518-1543 |
| [21] | Bandelt HJ, Dress AWM. Split decomposition: a new and useful approach to phylogenetic analysis of distance data[J]. Molecular Phylogenetics and Evolution, 1992, 1(3): 242-252 |
| [22] | Morrison DA. Networks in phylogenetic analysis: new tools for population biology[J]. International Journal for Parasitology, 2005, 35(5): 567-582 |
| [23] | Huson DH, Bryant D. Application of phylogenetic networks in evolutionary studies[J]. Molecular Biology and Evolution, 2006, 23(2): 254-267 |