 2015, Vol. 42
2015, Vol. 42扩展功能
文章信息
- 王茹, 郑平, 张萌, 赵和平, 周晓馨
- WANG Ru, ZHENG Ping, ZHANG Meng, ZHAO He-Ping, ZHOU Xiao-Xin
- 硝酸盐型厌氧铁氧化菌的种类、分布和特性
- Nitrate-dependentanaerobic ferrous/iron oxidation microorganism: review on its species,distribution and characteristics
- 微生物学通报, 2015, 42(12): 2448-2456
- Microbiology China, 2015, 42(12): 2448-2456
- 10.13344/j.microbiol.china.150200
-
文章历史
- 收稿日期: 2015-03-12
- 接受日期: 2015-05-25
- 优先数字出版日期(www.cnki.net): 2015-06-02

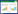




1996年,德国学者Straub等在溪流、城市沟渠等水体沉积物中观察到以硝酸盐为电子受体的铁氧化现象,称之为硝酸盐型厌氧铁氧化(Nitrate-dependent anaerobic ferrous/iron oxidation,NAFO);经过分离纯化,获得了具有NAFO能力的纯培养物,称之为硝酸盐型厌氧铁氧化菌(Nitrate-dependent anaerobic ferrous/iron oxidation microorganism,NAFOM)[1]。在此后20年的持续研究中发现,这类微生物广泛存在于自然界许多生境中,如淡水底泥、稻田土、海洋沉积物及地下水[2, 3, 4]。
NAFO的发现不仅为废水生物脱氮技术的开发提供了依据,也为NAFOM菌种的发掘和铁、氮循环的认识提供了新的线索[5, 6, 7]。本文整理了近20年文献报道的NAFOM资料,从系统分类学角度探讨了NAFOM的种类;从自然生境和人为生境两方面阐述了NAFOM的分布;从营养需求和生长代谢两层次分析了NAFOM的特性;从工程应用和生态系统铁、氮循环两个领域展示了NAFOM的研究意义。研究NAFOM的种类、分布和特性,了解NAFO研究进展,以期为NAFOM菌种资源的开发、地球铁素和氮素循环的研究、NAFO过程的优化提供借鉴。
1 NAFOM种类据文献报道,NAFOM广泛分布于细菌域和古菌域,涉及8个科,10个属(图 1)。在细菌域中,NAFOM主要分布于变形菌门,包括α-变形菌纲中1个科1个属、β-变形菌纲中3个科5个属及γ-变形菌纲中2个科2个属,同时包括放线菌纲中1个科1个属。在古菌域中,NAFOM仅见报道于广古菌门古丸菌纲古丸菌科铁丸菌属。

|
|
图 1
硝酸盐型厌氧铁氧化菌系统发育树
Figure 1
Phylogenetic tree of NAFOM
注:括号内为菌株NCBI序列号;分支点数字为步长1 000时百分比;标尺体现生物体与序列之间差异;分支长度对应序列与序列之间差异. Note: Serial numbers in brackets arefrom NCBI; Numbers at the nodes are percentages of bootstrap values based on 1 000re-samplings; The scale bar stands for the differences between organisms andsequence; Branch lengths correspond to sequence differences as indicated by thescale bar. |
NAFO细菌多呈杆状,大小为(1.5−2.5) μm× (0.5−1) μm,革兰氏阴性。NAFO古菌Ferroglobus placidus strain AEDII12DO为不规则球菌,表面有网格纹,直径0.7−1.3 μm,单生或对生,有1−2根鞭毛[8]。根据对氧气的需求状况,NAFOM可分为兼性厌氧菌和严格厌氧菌(表 1)。
| 种属 Species | 电子供体 Electron donor | 电子受体 Electron acceptor | 需氧型 Oxygen type | 营养型 Nutrition type | 生境 Habitat | 参考文献 Reference |
| Paracoccus ferrooxidans strain BDN-1 | [Fe(II)EDTA]2− | NO3−/NO2− | 兼氧 | 兼养 | 实验室反硝化生物流化床反应器 | [9] |
| Acidovorax sp. strain BoFeN1 | FeCl2 | NO3−/NO2− | 厌氧 | 兼养 | 德国康斯坦茨湖沿岸底泥沉积物 | [10, 11] |
| Acidovorax sp. strain 2AN | [Fe(II)EDTA]2− | NO3− | 厌氧 | 兼养 | 实验室缺氧序批式反应器 | [12, 13] |
| Acidovorax sp. strain BrG1 | FeSO4 | NO3−/NO2−/N2O | 兼氧 | 兼养 | 德国淡水沟渠、溪流和池塘底泥 | [1, 14, 15] |
| Aquabacterium sp. strain BrG2 | FeSO4 | NO3−/NO2−/N2O | 兼氧 | 兼养 | 德国淡水沟渠、溪流和池塘底泥 | [1, 14, 15] |
| Pseudogulbenkiania sp. strain 2002 | FeCl2 | NO3− | 厌氧 | 兼养 | 美国南伊利诺伊斯大学卡本代尔分校淡水湖底泥 | [16, 17] |
| Azospira sp. strain TR1 | FeSO4 | NO3− | 厌氧 | 自养 | 加拿大不列颠哥伦比亚省特雷尔某环境修复现场 | [18] |
| Dechloromonas sp. strain UWNR4 | [Fe(II)EDTA]2− | NO3− | 厌氧 | 兼养 | 美国威斯康星河河水底泥 | [10, 19] |
| Acidovorax sp. strain BrG3 | FeSO4 | NO3−/NO2− | 兼氧 | 兼养 | 德国淡水沟渠、溪流和池塘底泥 | [1, 14, 15] |
| Citrobacter freundii strain PXL1 | FeCl2 | NO3− | 厌氧 | 自养 | 乌鲁木齐河东污水处理厂 | [20] |
| Actinobacteria strain TM3 | FeCl2 | NO3− | 厌氧 | 自养 | 德国勃兰登堡梅克伦堡湖区底泥沉积物 | [21] |
| Ferroglobus placidus strain AEDII12DO | FeS | NO3− | 厌氧 | 自养 | 意大利火山岛附近的浅滩沉积物 | [8] |
NAFOM广泛分布于富含硝酸盐和亚铁盐的自然或人为生境中。
在自然生境中,已从封闭或半封闭淡水水源地地底泥样品内分离到不少NAFOM菌株(表 1)。这类生境与水体的物质交流较少,生态条件相对稳定,缺氧但富含硝酸盐和亚铁盐,适合NAFOM生活;由于处于淡水环境中,这类NAFOM大多不耐盐(<4.5% NaCl)。Actinobacteria strain TM3分离自德国勃兰登堡梅克伦堡湖区GrosseFuchskuhle湖,因接纳临近沼泽地的部分沼液,湖水呈酸性(pH 4.5),底泥中富含腐殖酸、硝酸盐(2 μmol/L)和亚铁盐(3−40 μmol/L)[21]。
在人为生境中,已从废弃物处理系统的活性污泥样品内分离到NAFOM菌株。Paracoccus ferrooxidans strain BDN-1分离自生物脱氮流化床反应器[9];Acidovorax sp. strain 2AN分离自序批式生物铁氧化反应器[11]。Azospira sp. strain TR1分离自某工业垃圾填埋场渗滤液污染环境修复现场[18]。
迄今为止,文献报道的硝酸盐型厌氧铁氧化古菌只有1株,即Archaeoglobacea ferroglobus placidusstrain AEDII12DO,它分离于意大利火山岛附近的浅滩沉积物中,沉积物温度为95 °C,pH为7.0,铁氧化物含量丰富[8]。
3 NAFOM特性 3.1 营养特性就能源需求而言,NAFOM属化能营养型。自养NAFOM通过氧化二价铁获取能量;兼养NAFOM通过氧化二价铁或有机物获取能量。
就碳源需求而言,NAFOM属兼性营养型。自养NAFOM以碳酸氢盐[20]或二氧化碳[8]为碳源;兼养NAFOM碳源广泛,包括以甲醇、乳糖等为代表的有机物[16]和以碳酸氢盐、二氧化碳等为代表的无机物[10]。
就氮源需求而言,NAFOM可利用硝酸盐、亚硝酸盐等无机态氮源,少部分可利用一氧化二氮为氮源[1, 14];多数NAFOM还可利用组氨酸、丝氨酸和天冬氨酸等有机态氮源[12, 13]。
部分兼养NAFOM的有机物利用性能见表 2。
| 有机物 Items | 菌株 Strains | ||||||
| BrG1 | BrG2 | BrG3 | 2AN | BDN1 | BoFeN1 | 2002 | |
| 乙酸盐 Acetate | + | + | + | NM | + | + | NM |
| 丙酸盐 Propionate | + | + | ‒ | NM | + | + | NM |
| 丁酸盐 Butyrate | + | + | ‒ | NM | NM | + | NM |
| 苹果酸盐 Malate | + | + | ‒ | + | NM | + | NM |
| 柠檬酸盐 Citrate | + | ‒ | ‒ | + | ‒ | ‒ | NM |
| 延胡索酸盐 Fumarate | + | + | ‒ | + | NM | + | NM |
| 琥珀酸盐 Succinate | + | + | ‒ | NM | + | + | NM |
| 甲醇 Methanol | NM | NM | NM | NM | + | NM | NM |
| 乙醇 Ethanol | + | + | ‒ | NM | NM | + | NM |
| 正丙醇 1-Propanol | + | + | ‒ | NM | NM | + | NM |
| 葡萄糖 Glucose | ‒ | ‒ | + | NM | + | ‒ | + |
| 果糖 Fructose | ‒ | ‒ | NM | NM | + | ‒ | NM |
| 乳糖 Lactose | NM | NM | NM | NM | ‒ | NM | NM |
| 麦芽糖 Maltose | ‒ | ‒ | + | NM | NM | + | |
| 丝氨酸 Serine | ‒ | ‒ | + | NM | + | NM | NM |
| 组氨酸 Histidine | NM | NM | NM | NM | + | NM | NM |
| 天门冬氨酸 Aspartate | + | ‒ | + | NM | NM | NM | NM |
| 谷氨酸盐 Glutamate | + | ‒ | + | NM | NM | NM | NM |
| 甘油 Glycerinum | NM | NM | NM | NM | NM | ‒ | NM |
NAFOM将2价铁氧化成3价铁,后者以致密氧化物或氢氧化物形式聚集在细胞表面[22]。图 2所示为本课题获得的NAFOM表面覆盖铁沉积物照片。胞外堆积的铁氧化物或氢氧化铁可阻碍细胞与外界的物质交流以及细胞与细胞之间的信息交流,从而影响NAFOM生长和NAFO活性。

|
| 图 2 2NAFOM扫描电镜照片 Figure 2 Scanning electron micrographs of NAFOM |
为维持生长,NAFOM可去除细胞表面的3价铁,其机制有多种:(1) 通过提高细胞表面正电荷数,与胞外3价铁形成排斥作用,使3价铁远离细胞壁,避免3价铁在细胞表面形成固态沉积物[22](图 3a);(2)通过质子泵泵出质子,产生微酸性环境,使近细胞壁的3价铁呈溶解态,当3价铁远离细胞壁时,在中性溶液中形成3价铁氧化物或氢氧化物沉淀[23, 24](图 3b);(3) 释放有机配体,与3价铁形成可溶性复合物,避免3价铁沉积[25](图 3c)。

|
| 图 3 NAFOM对Fe3+的迁移机理 Figure 3 Fe3+ transportionmechanism by NAFOM |
NAFO过程的电子供体为Fe2+,其化学形态有[Fe(II)EDTA]2−、FeCO3、FeCl2、FeS和FeSO4等;电子受体为NO3−和NO2−,其化学形态有NaNO3、KNO3、NaNO2和KNO2等。
根据氧化还原电位(图 4),NO3−/½N2的标准电极电位高出[Fe(OH)3/Fe2+](0.699 V),NO2−/½N2的标准电极电位高出[Fe(OH)3/Fe2+](0.268 V)。从电对势能看,NAFO过程为正向电子传递,微生物可从NAFO过程获取能量。

|
| 图 4 电子供体与电子受体的氧还电位 Figure 4 Redox potential electron donors and acceptors |
亚铁型反硝化反应的电极电位较之乙酸型异养反硝化低0.267 V,较之氢气型、甲烷型、硫化物型自养反硝化分别低0.400、0.238、0.210 V。而铁型反硝化则具有强大电极电位优势,较之乙酸型异养反硝化高0.152 V,较之氢气型、甲烷型、硫化物型自养反硝化分别高0.019、0.181、0.209 V。就电极电位角度而言,铁型反硝化过程更易发生。
| $2N{{O}_{3}}^{-}+10F{{e}^{2+}}+24{{H}_{2}}O\to 10Fe{{(OH)}_{3}}+{{N}_{2}}+18{{H}^{+}}\ \ \ \ {{\Delta }_{r}}G_{m}^{\theta }=-472.72\text{ }kJ\cdot mol$ | (1) |
| $2N{{O}_{3}}^{-}+10F{{e}^{2+}}+24{{H}_{2}}O\to 10Fe{{(OH)}_{3}}+{{N}_{2}}+18{{H}^{+}}\ \ \ \ {{\Delta }_{r}}G_{m}^{\theta }=-472.72\text{ }kJ\cdot mol$ | (2) |
根据化学反应方程式(1)和(2),反应物NO3−/Fe2+(M/M)为1:5,NO2−/Fe2+(M/M)为1:3。反应产物多为氢氧化铁和氮气。反应吉布斯自由能小于零,化学反应能自发发生,产生的能量可供微生物代谢。
表 3列出部分NAFOM的NAFO活性。对于兼性化能自养NAFOM,添加有机物可促进NAFO活性,例如,添加乙酸盐(≤1 mmol/L),Acidovorax sp. strain BrG1的亚铁转化速率提升4倍[1];添加富马酸(≤0.5 mmol/L),Aquabacterium sp. strain BrG2的亚铁转化速率提升3倍[1];添加0.5−1.0 mmol/L乙酸盐,Acidovorax sp. strain 2AN可长效、连续进行NAFO反应[12]。
| 种属 Species | 硝氮浓度 Nitrate concentration (mmol/L) | 亚铁浓度 Ferrous concentration (mmol/L) | 反应时间 Reaction time (d) | 硝氮去除 Nitrate removal (%) | 亚铁去除Ferrous removal (%) | 有机物添加Organic added (mmol/L) |
| Acidovorax sp. strain BrG1 | 4.0 | 10.0 | 8 | 50.0 | 58.0 | 0.5乙酸盐 |
| Aquabacterium sp. strain BrG2 | 4.0 | 10.0 | 21 | 100.0 | 83.0 | 无有机物添加 |
| Dechloromonas sp. strain UWNR4 | 4.7 | 6.1 | 3 | 51.1 | 88.5 | 1.2乙酸盐 |
| Acidovorax sp. strain 2AN | 5.0 | 8.0 | 7 | 43.8 | 90.0 | 1.6乙酸盐 |
| Paracoccus ferrooxidans strain BDN-1 | 5.0 | 25.0 | 2−3 | NM | 100.0 | 1乙醇 |
| Acidovorax sp. strain BoFeN1 | 5.0 | 2.0 | 6 | 100.0 | 100.0 | 2乙酸盐 |
| Pseudogulbenkiania sp. strain 2002 | 2.3 | 10.0 | 7 | 16.1 | 24.5 | 0.1乙酸盐 |
| Azospira sp. strain TR1 | 4.0 | 10.0 | 18 | NM | 70.0 | 无有机物添加 |
| Citrobacter freundii sp. strain PXL1 | 9.5 | 3.4 | 3 | 30.0 | 85.0 | 无有机物添加 |
有关有机物对NAFOM活性的刺激效应,2009年Weber等提出了“共基质”的解释[17]。有机物充足时,有机物作为“第一基质”,为NAFOM生长提供碳源及能源,亚铁作为“第二基质”,为微生物生长提供能源,丰富的碳源和能源有利于NAFOM生长及NAFO活性提高;有机物匮乏时,NAFOM只能氧化亚铁获得能量,NAFOM生长缓慢,NAFO活性降低。
4 NAFOM应用 4.1 废水处理与传统异养反硝化过程相比,NAFO利用低价铁作为电子供体进行反硝化,具有经济成本低、二次污染少、且不产生温室气体等优势。将NAFO作为一种新型技术应用于废水脱氮处理,具有工程价值和应用前景。NAFO工艺处于研发初期,目前只见实验室NAFO小试报道。张萌等还以造纸厂IC反应器厌氧消化污泥作为接种源,采用模拟废水运行NAFO反应器,取得良好成效,稳定运行时硝氮和亚铁的去除率分别为45.63%±3.31%和43.94%±3.55%,容积负荷和容积去除速率分别为0.159±0.01 kg N/(m3·d)和0.073±0.01 kg N/(m3·d)[26]。
此外,学者还研究了异养反硝化菌和厌氧氨氧化菌参与的NAFO反应。Wang等首次报道了异养反硝化活性污泥具有NAFO活性[27],Zhang等研究了理化性质,如Fe/N、温度以及起始pH等,对该反硝化颗粒污泥NAFO活性的影响[28]。Oshiki等研究了厌氧氨氧化菌的NAFO活性,厌氧氨氧化菌Candidatus brocadia sinica富集培养物对亚铁的氧化速率为3.7±1.2 μmol/(gprotein·min),对硝氮的还原速率为1.3±0.1 μmol/(g protein·min)[29]。
NAFO作为一种新型废水脱氮技术,不仅实现废水自养脱氮目的,其反应产物三价铁还可作为高效废水除磷絮凝剂,同步实现废水脱氮除磷。针对含氮含磷废水,如生活污水和养殖废水等,研发NAFO技术,将大大缩减处理成本,具有非常可观的经济价值。
4.2 生态研究地壳中铁元素含量丰富,广泛存在于淡水、海水及各种矿物中,铁氧化菌和铁还原菌参与的地壳铁循环已逐步为人们所熟识[30, 31]。氮化合物种类多样,包括呈溶解态的硝酸盐和亚硝酸盐,和呈气态的氮气和氮氧化物,微生物参与的地壳氮循环也已探究透彻[32, 33]。传统意义上讲,铁循环和氮循环被认为是两个独立的过程,并无交集。然而,NAFO的发现却让我们认识到一种新的地壳铁氮循环方式。NAFOM通过氧化亚铁获取生存生长能量,以硝酸盐作为电子受体进行厌氧呼吸,其在自然界氮、铁循环中发挥至关重要作用,研究NAFOM有助于进一步认识地球物质循环,探明NAFOM代谢机理有助于了解古生物耦合硝酸盐还原和亚铁氧化的过程特性[34, 35]。
此外,研究NAFOM还有助于探明前寒武世纪铁地质层形成机理,有助于探索火星等铁矿丰富的外星球上是否存在生命现象[36, 37]。
5 小结与展望NAFOM是以低价铁作为电子供体的反硝化作用,它是微生物领域的重大发现。研究NAFOM有助于发掘反硝化新种,丰富反硝化菌种资源库;有助于探明微生物氮还原和铁氧化的代谢途径,丰富微生物生理学知识;有助于了解地球铁、氮循环及微生物进化史;有助于开发新型废水脱氮技术,促进污染环境的生物修复。
然而,目前国内外对NAFOM的研究还处于起步阶段,国外学者多围绕NAFOM参与的地球铁、氮循环展开研究,未涉及NAFOM的代谢机理及水处理工程应用;国内学者多围绕铁盐耦合反硝化进行废水脱氮技术研究,未曾对NAFOM的铁盐脱氮机理及反应条件优化等方面进行探索[38, 39, 40]。笔者认为,NAFO虽具有巨大的潜在应用价值,但离工程应用还有很大差距。今后研究工作的重点应放在NAFO高效菌株的分离筛选及生长代谢特性研究,NAFO过程机理及反应条件优化,NAFO小试装置的设计研发及运行调试等。
| [1] | Straub KL, Benz M, Schink B, et al. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron[J].Applied and Environmental Microbiology, 1996, 62(4): 1458-1460 |
| [2] | Hauck S, Benz M, Brune A, et al. Ferrous iron oxidation by denitrifying bacteria in profundal sediments of a deep lake (Lake Constance)[J].FEMS Microbiology Ecology, 2001, 37(2): 127-134 |
| [3] | Hegler F, Losekann-Behrens T, Hanselmann K, et al. Influence of seasonal and geochemical changes on the geomicrobiology of an iron carbonate mineral water spring[J].Applied Environmental Microbiology, 2012, 78(20): 7185-7196 |
| [4] | Jorgensen CJ, Jacobsen OS, Elberling B, et al. Microbial oxidation of pyrite coupled to nitrate reduction in anoxic groundwater sediment[J].Environmental Science Technology, 2009, 43(13): 4851-4857 |
| [5] | Zhang M, Zheng P, Ji JY. Research advances on anaerobic ferrous-oxidizing microorganisms[J].Journal of Applied Ecology, 2013, 24(8): 2377-2382 (in Chinese)张萌, 郑平, 季军远. 厌氧铁氧化菌研究进展[J].应用生态学报, 2013, 24(8): 2377-2382 |
| [6] | Zhang M, Zheng P. Clarification, habitat and enrichment technique of Fe(II)- oxidizing microorganisms[J].Bulletin of Science and Technology, 2012, 28(11): 72-76,152(in Chinese)张萌, 郑平. 铁氧化菌的类型、生境及富集培养方法[J].科技通报, 2012, 28(11): 72-76,152 |
| [7] | Wang HY, Zhang HN, Lü B, et al. Research on nitrate removal by Fe-dependent autotrophic denitrification bacteria[J].China Ryral Water and Hydropower, 2014(11): 59-62 (in Chinese)王弘宇, 张惠宁, 吕斌, 等. 铁基质自养反硝化去除水中硝酸盐污染物的研究[J].中国农村水利水电, 2014(11): 59-62 |
| [8] | Hafenbradl D, Keller M, Dirmeier R, et al. Ferroglobus placidus gen. nov., sp. nov., a novel hyperthermophilic archaeum that oxidizes Fe2+ at neutral pH under anoxic conditions[J].Archives of Microbiology, 1996, 166(5): 308-314 |
| [9] | Kumaraswamy R, Sjollema K, Kuenen G, et al. Nitrate- dependent[Fe(II)EDTA]2? oxidation by Paracoccus ferrooxidans sp. nov., isolated from a denitrifying bioreactor[J].Systematic and Applied Microbiology, 2006, 29(4): 276-286 |
| [10] | Kappler A, Schink B, Newman DK. Fe(III) mineral formation and cell encrustation by the nitrate-dependent Fe(II)-oxidizer strain BoFeN1[J].Geobiology, 2005, 3(4): 235-245 |
| [11] | Miot J, Benzerara K, Morin G, et al. Iron biomineralization by anaerobic neutrophilic iron- oxidizing bacteria[J].Geochimica et Cosmochimica Acta, 2009, 73(3): 696-711 |
| [12] | Chakraborty A, Roden EE, Schieber J, et al. Enhanced growth of Acidovorax sp. strain 2AN during nitrate- dependent Fe(II) oxidation in batch and continuous-flow systems[J].Applied and Environmental Microbiology, 2011,77(24): 8548-8556 |
| [13] | Chakraborty A, Picardal F. Induction of nitrate-dependent Fe(II) oxidation by Fe(II) in Dechloromonas sp. strain UWNR4 and Acidovorax sp. strain 2AN[J].Applied and Environmental Microbiology, 2013, 79(2): 748-752 |
| [14] | Straub KL, Buchholz-Cleven BEE. Enumeration and detection of anaerobic ferrous iron-oxidizing, nitrate- reducing bacteria from diverse European sediments[J].Applied and Environmental Microbiology, 1998, 64(12): 4846-4856 |
| [15] | Straub KL, Sch?nhuber WA, Buchholz-Cleven BE, et al. Diversity of ferrous iron-oxidizing, nitrate-reducing bacteria and their involvement in oxygen-independent iron cycling[J].Geomicrobiology Journal, 2004, 21(6): 371-378 |
| [16] | Weber KA, Pollock J, Cole KA, et al. Anaerobic nitrate-dependent iron (II) bio-oxidation by a novel lithoautotrophic betaproteobacterium, strain 2002[J].Applied and Environmental Microbiology, 2006, 72(1): 686-694 |
| [17] | Weber KA, Hedrick DB, Peacock AD, et al. Physiological and taxonomic description of the novel autotrophic, metal oxidizing bacterium, Pseudogulbenkiania sp. strain 2002[J].Applied Microbiology and Biotechnology, 2009, 83(3): 555-565 |
| [18] | Mattes A, Gould D, Taupp M, et al. A novel autotrophic bacterium isolated from an engineered wetland system links nitrate-coupled iron oxidation to the removal of As, Zn and S[J].Water, Air, & Soil Pollution, 2013, 224(4): 1-15 |
| [19] | Chakraborty A, Picardal F. Neutrophilic, nitrate-dependent, Fe(II) oxidation by a Dechloromonas species[J].World Journal of Microbiol Biotechnol, 2013, 29(4): 617-623 |
| [20] | Li B, Tian C, Zhang D, et al. Anaerobic nitrate-dependent iron (II) oxidation by a novel autotrophic bacterium, Citrobacter freundii strain PXL1[J].Geomicrobiology Journal, 2014, 31(2): 138-144 |
| [21] | Kanaparthi D, Pommerenke B, Casper P, et al. Chemolithotrophic nitrate-dependent Fe(II)-oxidizing nature of actinobacterial subdivision lineage TM3[J].ISME Journal, 2013, 7(8): 1582-94 |
| [22] | Sch?dler S, Burkhardt C, Hegler F, et al. Formation of cell-iron-mineral aggregates by phototrophic and nitrate- reducing anaerobic Fe(II)-oxidizing bacteria[J].Geomicrobiology Journal, 2009, 26(2): 93-103 |
| [23] | Urrutia MM, Kemper M, Doyle R, et al. The membrane-induced proton motive force influences the metal binding ability of Bacillus subtilis cell walls[J].Applied Environmental and Microbiology, 1992, 58(12): 3837-3844 |
| [24] | Johnson KJ, Ams DA, Wedel AN, et al. The impact of metabolic state on Cd adsorption onto bacterial cells[J].Geobiology, 2007, 5(3): 211-218 |
| [25] | Kappler A, Newman DK. Formation of Fe(III)-Minerals by Fe(II)-oxidizing photoautotrophic bacteria[J].Geochimica et Cosmochimica Acta, 2004, 68(6): 1217-1226 |
| [26] | Zhang M, Zheng P, Li W, et al. Performance of nitrate-dependent anaerobic ferrous oxidizing (NAFO) process: A novel prospective technology for autotrophic denitrification[J].Bioresource Technology, 2015, 179: 543-548 |
| [27] | Wang R, Zheng P, Xing YJ, et al. Anaerobic ferrous oxidation by heterotrophic denitrifying enriched culture[J].Journal of Industrial Microbiology and Biotechnology, 2014, 41(5): 803-809 |
| [28] | Zhang M, Zheng P, Wang R, et al. Nitrate-dependent anaerobic ferrous oxidation (NAFO) by denitrifying bacteria: a perspective autotrophic nitrogen pollution control technology[J].Chemosphere, 2014, 117: 604-609 |
| [29] | Oshiki M, Ishii S, Yoshida K, et al. Nitrate-dependent ferrous iron oxidation by anaerobic ammonium oxidation (anammox) bacteria[J].Applied and Environmental Microbiology, 2013, 79(13): 4087-4093 |
| [30] | Hedrich S, Schl?mann M, Johnson DB. The iron-oxidizing proteobacteria[J].Microbiology, 2011, 157(6): 1551-1564 |
| [31] | Kappler A, Straub KL. Geomicrobiological cycling of iron[J].Molecular Geomicrobiology, 2005, 59(1): 85-108 |
| [32] | Sparacino-Watkins C, Stolz JF, Basu P. Nitrate and periplasmic nitrate reductases[J].Chemical Society Reviews, 2014, 43(2): 676-706 |
| [33] | Zumft WG. Cell biology and molecular basis of denitrification[J].Microbiology and Molecular Biology Reviews, 1997, 61(4): 533-616 |
| [34] | Weber KA, Achenbach LA, Coates JD. Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction[J].Nature Reviews Microbiology, 2006, 4: 752-764 |
| [35] | Muehe EM, Gerhardt S, Schink B, et al. Ecophysiology and the energetic benefit of mixotrophic Fe(II) oxidation by various strains of nitrate-reducing bacteria[J].FEMS Microbiology Ecology, 2009, 70(3): 335-343 |
| [36] | Ghiorse WC. Biology of iron-and manganese-depositing bacteria[J].Annual Review of Microbiology, 1984, 38: 515-550 |
| [37] | Widdel F, Schnell S, Heising S, et al. Ferrous iron oxidation by anoxygenic phototrophic bacteria[J].Nature, 1993, 362(6423): 834-836 |
| [38] | Feng H, Liang YX, Du Y, et al. Effect of microbial redox cycling of iron on denitrification in landfill[J].Acta Scientiae Circumstantiae, 2014, 34(2): 409-416 (in Chinese)冯欢, 梁禹翔, 杜耀, 等. 填埋场中铁的生物化学循环对反硝化的影响[J].环境科学学报, 2014, 34(2): 409-416 |
| [39] | Shen DS, Li WB, Yao J, et al. Microbial-mediated anaerobic oxidation of fe rrous iron and its mech anism to remediate contaminated environments[J].Journal of Zhejiang University, 2011, 37(1): 112-118 (in Chinese)沈东升, 李文兵, 姚俊, 等. 亚铁厌氧微生物氧化及其在环境污染修复中的作用机制[J].浙江大学学报, 2011, 37(1): 112-118 |
| [40] | Wang X, Li TL, Dong MY, et al. Reducing nitrate in groundwater by stabilized iron nanoparticles and denitrifying bacteria[J].Journal of Agro-Environment Science, 2011, 30(4): 739-745 (in Chinese)王学, 李铁龙, 东美英, 等. 稳定纳米铁与反硝化菌耦合去除地下水中硝酸盐的研究[J].农业环境科学学报, 2011, 30(4): 739-745 |