 2015, Vol. 42
2015, Vol. 42扩展功能
文章信息
- 刘蒙蒙, 宋超, 邢咏梅, 郭顺星
- LIU Meng-Meng, SONG Chao, XING Yong-Mei, GUO Shun-Xing
- 猪苓菌丝形成菌核的MAPK基因克隆及表达分析
- Molecular cloning and characterization of a MAPK gene from Polyporus umbellatus
- 微生物学通报, 2015, 42(12): 2345-2350
- Microbiology China, 2015, 42(12): 2345-2350
- 10.13344/j.microbiol.china.150150
-
文章历史
- 收稿日期: 2015-02-17
- 接受日期: 2015-04-15
- 优先数字出版日期(www.cnki.net): 2015-05-08

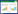




丝裂原活化蛋白激酶(Mitogen-activated protein kinases,MAPKs)是细胞内的一类丝氨酸/苏氨酸蛋白激酶[1]。研究证实,MAPKs信号转导通路存在于大多数细胞内,在将细胞外刺激信号转导至细胞及其核内,并引起细胞生物学反应(如细胞增殖、分化、转化及凋亡等)的过程中具有至关重要的作用。研究表明,MAPKs信号转导通路在细胞内具有生物进化的高度保守性,在低等原核细胞和高等哺乳类细胞内,目前均已发现存在着多条并行的MAPKs信号通路,不同的细胞外刺激可使用不同的MAPKs信号通路,通过其相互调控而介导不同的细胞生物学反应[2]。
真菌中共发现了22个MAPK基因,根据这些MAPK基因的功能,可将其分为以下4类:与真菌生长发育相关的基因[3]、与致病性有关的基因、与胁迫反应相关的基因[4]以及功能未知的基因。参与真菌生长发育的MAPK基因主要参与调节菌丝发育、分生孢子或子囊孢子的形成、萌发等生理过程。到目前为止,对于大型真菌子实体形成以及菌核发育的MAPK基因缺少相应的研究。
猪苓是我国重要的药用真菌,因其菌核具有良好的利水渗湿药用价值而受到广泛的关注。由于对野生猪苓资源毫无节制的采挖,造成其资源的日益匮乏。猪苓菌丝形成菌核是解决猪苓资源匮乏的主要途径之一。目前对猪苓的研究已在中药化学[5]、药理学[6]、解剖生物学及生理生化[7]等方面取得了诸多成就。但对猪苓菌丝形成菌核发育的分子机制缺少研究。本研究利用RACE技术从猪苓菌核中分离到一个MAPK基因,并对其进行生物信息学及猪苓菌丝形成菌核过程中表达模式分析,为进一步揭示其在猪苓菌核生长发育过程中的生物学功能提供依据。
1 材料与方法 1.1 供试菌株猪苓纯培养菌丝由中国医学科学院药用植物研究所真菌室提供,该菌种由本实验室保藏,编号为ZL-001。
1.2 猪苓菌核培养依据本实验室报道的方法[8]将猪苓菌丝接种到人工培养基中。培养20−30 d后,猪苓菌丝形成菌核,30−40 d为猪苓菌核的快速生长时期。分别将初期菌核、发育菌核(SI、SD)和生长时期未形成菌核的菌丝(HI、HD)立即放入液氮中预冷并−80 °C保存以备提取RNA。
1.3 总RNA提取及质量控制利用EASYspin Plus Kit (Aidlab,中国)试剂盒并根据操作手册进行总RNA的提取。利用凝胶电泳检测RNA的完整性并通过NanoDropTM 2000分光光度计(Thermo Fisher Scientific,美国)检测样品总RNA的质量和纯度。
1.4 5′-RACE-PCR和基因克隆为了获得完整的MAPK基因全长序列,采用5′-RACE-PCR方法获取猪苓MAPK全长cDNA。根据猪苓EST序列Pu1317使用Primer5软件设计5′引物GSP1 (5′-GCCAGAGGATTTGCGTTCGGG AAG-3′)。5′-RACE实验使用SMART™ RACE cDNA Amplification试剂盒(Clontech,美国)。
利用BD PowerScriptTM Reverse Transcriptase (Clotech,美国)试剂盒获取5′-RACE-Ready cDNA。分别以GSP1与试剂盒提供的UPM引物组合进行5′-RACE。反应体系:10×Advantage® 2 PCR buffer 2.5 µL,10 mmol/L dNTPs 0.5 µL,10 µmol/L GSP1 0.5 µL,10×UPM (Universal Primer A Mix) 0.5 µL,5-RACE-Ready cDNA 1 µL,50×Advatange® 2 Polymerase Mix (5 U/L) 0.5 µL,19.5 µL ddH2O,总反应体积为25 µL。反应条件:94 °C 30 s,72 °C 3 min,5个循环;94 °C 30 s,70 °C 30 s,72 °C 3 min,5个循环;94 °C 30 s,68 °C 30 s,72 °C 3 min,20个循环;72 °C 7 min。PCR反应产物经2%琼脂糖凝胶电泳分离后得到目的条带,经切胶纯化后克隆到pGEM-T载体(Promega,美国),然后转化入E. coli JM109细胞(TaKaRa,中国)中。阳性克隆由北京金唯智生物科技有限公司进行测序。
1.5 序列分析使用多种在线生物信息学分析工具对得到的全长cDNA进行序列分析:BLAST (http://www.ncbi.nlm.nih.gov/blast/),ORF Finder (http://www.ncbi.nlh.nih.gov/gorf/gorf.html),以及CAP3 Sequence Assembly Program (http://pbil.univ-lyon1.fr/cap3.php/)。应用DNAstar 6.0 (Madison,美国)进行多序列比对。用MEGA (Version 5.0)软件对MAPK进行系统发育分析和进化树的构建。进化距离分析采用邻接法(Neighbor-Joining,NJ)。
1.6 实时荧光定量PCR分析利用MMLV Reverse Transcriptase (Promega,美国)试剂盒并依据操作手册对样本SI、SD、HI和HD进行反转录。利用实时荧光定量PCR方法分析不同样本中猪苓MAPK基因的表达量变化,以猪苓18S rRNA 基因作为内参基因。根据已知的猪苓MAPK基因EST序列设计实时荧光定量PCR引物:qPCR-FP (5′-ATTCGTGCGCTGCCGTTC-3′)和qPCR-RP (5′-GATGCGCTTCTTCGGGTCA-3′)。qPCR反应按照SYBR® Premix ExTaqTM试剂盒(TaKaRa,中国)的操作手册进行。PCR反应体系(25 µL):2×SYBR® Premix ExTaqTM Master Mix 12.5 μL,引物(10 μmol/L)各0.5 μL,ROX Reference Dye 0.5 μL,cDNA 模板2 μL,ddH2O 9 μL。反应条件:95 °C 30 s;95 °C 5 s,60 °C 34 s,40个循环。同时做熔解曲线。每个样本的qPCR反应做3次重复实验。Ct (Cycle threshold)值由ABI PRISM 7500软件生成,采用∆∆CT法计算相对表达量[9]。
2 结果与分析 2.1 猪苓MAPK基因的克隆由于原EST序列具有完整的3′端,以GSP1为引物,通过5′-RACE-PCR反应,克隆、测序获得827 bp的序列,与原来的EST序列拼接分析获得了一条1 293 bp的猪苓MAPK全长cDNA。BLASTx比对分析结果显示其与Postia placenta MAPK C端氨基酸残基具有97%一致性。因此将其定名为PuMAPK,提交GenBank获得注册号JX035916。根据基因全长cDNA推定其氨基酸序列。猪苓MAPK (PuMAPK)基因的开放阅读框(Open reading frame,ORF)长1 161 bp,编码386个氨基酸,推测分子量为43.872 kD,理论等电点为6.68。
2.2 MAPK基因编码蛋白的多序列比对分析在NCBI对猪苓MAPK进行BLASTp分析(图 1),结果所示,PuMAPK有MAPK中ERK1/2类型的保守区。运用DNAStar 6.0中的MegAlign程序,猪苓PuMAPK推定的氨基酸序列和其他6种真菌MAPK蛋白进行多序列比对分析(图 2)。结果表明,PuMAPK与其他来源于担子菌的MAPK序列基本一致(图 2)。PuMAPK与Coprinus cinereus、Laccaria bicolor、Volvariella volvacea、Postia placenta、Piriformospora indica的MAPK序列一致性分别高达96%、95%、93%、97%和95%,与核盘菌(Sclerotinia sclerotiorum)的SMK1一致性为73%。且包括PuMAPK在内的这类MAPK具有TEY结构位点。分析结果表明本研究克隆得到的PuMAPK其推定的氨基酸序列具有MAPK结构域,PuMAPK为一新的编码MAPK基因。

|
| 图 1 Blastp分析猪苓PuMAPK基因全长cDNA的结构域 Figure 1 Analyses of PuMAPK cDNA domains of by Blastp |

|
| 图 2 猪苓MAPK蛋白序列(Pu1317)与担子菌MAPK蛋白的多序列比对 Figure 2 Multiple sequence alignment of PuMAPK and MAPK proteins from other fungus |
通过NCBI检索选取高等真菌中的MAPK序列利用MEGA 5.0构建了包括PuMAPK (Pu1317)在内共17个MAPK的系统进化树(图 3)。序列包括Coprinus cinereus、Laccaria bicolor、Volvariella volvacea、Postia placenta、Piriformospora indica、Puccinia triticina、Sclerotinia sclerotiorum、Claviceps purpurea、Neurospora crass、Cryphonectria parasitica、Trichoderma reesei、Nectria haematococca、Fusarium oxysporum、Mus musculus、Saccharomyces cerevisiae和Cryptococcus neoformans。Schizosaccharomyces cerevisiae和Cryptococcus neoformans形成第一个分支,支持度为100。担子菌群和子囊菌群聚类在两个姊妹群系中,支持度为100。PuMAPK与担子菌类群聚集在一起,说明其与担子菌亲缘关系较近。

|
| 图 3 猪苓MAPK基因序列的系统发育树 Figure 3 Phylogenetic tree of PuMAPK with various MAPK proteins Note:Numbers in parentheses represent the sequences’ accession number in GenBank. The number at each branch points is the percentage supported by bootstrap. Bar, 5% sequence divergence. |
分别取原基期(SI)和生长期(SD)的菌核和未形成菌核的菌丝体(HI、HD)总RNA,利用荧光定量PCR方法测定mRNA的表达量。以菌丝样本为对照组,检测了猪苓PuMAPK在不同生长时期菌核和菌丝组织间的转录水平差异(图 4)。菌核形成初期PuMAPK在菌核中表达量高于菌丝7.863倍,至生长期时在菌核中的表达量仅为菌丝1.26倍。结果表明在猪苓菌核形成初期,菌核中的MAPK表达量显著高于菌丝组织,随着菌核的快速生长而减少。

|
| 图 4 菌核形成过程中猪苓MAPK的差异表达 Figure 4 Differential expression analysis of MAPK gene during sclerotial growth |
丝裂原活化蛋白激酶是信号从细胞表面传导到细胞核内部的重要传递者。在真菌中,并行的MAPKs信号通路在细胞信号转导中并无相互作用,其每一条MAPKs通路都是相对独立的,通常不与其他通路发生交联[4]。对真菌而言,不同的MAPKs通路调节不同的生理过程;对于同样的刺激,几条并行的通路并不同时被激活;其中一条通路若出现突变,也不影响其他通路的信号传递。细胞外信号调节蛋白激酶(Extracellular signal regulated protein kinases,ERKs)是MAPK家族中的3个成员之一[10]。ERKs有ERK1和ERK2两种同工酶。一般认为,ERK主要在生长因子相关刺激引起的细胞反应中起作用,主要参与细胞生长、分化和抗凋亡作用[11]。H2O2预处理能持续激活ERK1/2,ERK活化能对抗氧化应激对细胞存活的损伤,ERK通路是适应性细胞保护机制之一。Chen等[12]在核盘菌中鉴定得到一个与ERK同源的smk1,该基因转录水平和其编码的MAPK蛋白活性在菌核发育,尤其是菌核形成初期显著提高。当添加MAPK抑制剂时,smk1表达受阻,菌核发现受抑制。筛选获得smk1突变体,该突变体不能形成菌核。研究结果表明,核盘菌菌核形成发育受ERK类型MAPK的调控。本研究在猪苓中同样克隆得到与smk1一致性为73%的ERK类型MAPK基因PuMAPK,且该基因在菌核形成初期时在菌核组织中高水平转录,表明该基因可能调控了猪苓菌核的形成。本研究将进一步通过RNAi研究PuMAPK基因在猪苓菌核生长发育中的生物学功能,为猪苓菌丝形成菌核的研究提供分子机制的理论基础。
| [1] | Darling NJ, Cook SJ. The role of MAPK signalling pathways in the response to endoplasmic reticulum stress[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2014, 1843(10): 2150-2163 |
| [2] | Pitzschke A. Modes of MAPK substrate recognition and control[J]. Trends in Plant Science, 2015, 20(1): 49-55 |
| [3] | Kitade Y, Sumita T, Izumitsu K, et al. MAPKK-encoding gene Ste7 in Bipolaris maydis is required for development and morphogenesis[J]. Mycoscience, 2015, 56(2): 150-158 |
| [4] | Alonso-Monge R, Román E, Arana DM, et al. Fungi sensing environmental stress[J]. Clinical Microbiology and Infection, 2009, 15(S1): 17-19 |
| [5] | Zhao YY, Cheng XL, Zhang Y, et al. Simultaneous determination of eight major steroids from Polyporus umbellatus by high-performance liquid chromatography coupled with mass spectrometry detection[J]. Biomedical Chromatography, 2010, 24(2): 222-230 |
| [6] | Zhao YY, Xie RM, Chao X, et al. Bioactivity-directed isolation, identification of diuretic compounds from Polyporus umbellatus[J]. Journal of Ethnopharmacology, 2009, 126(1): 184-187 |
| [7] | Xing YM, Chen J, Lü YL, et al. Determination of optimal carbon source and pH value for sclerotial formation of Polyporus umbellatus under artificial conditions[J]. Mycological Progress, 2011, 10(1): 121-125 |
| [8] | Liu YY, Guo SX. Nutritional factors determining sclerotial formation of Polyporus umbellatus[J]. Letters in Applied Microbiology, 2009, 49(2): 283-288 |
| [9] | Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR[J]. Nucleic Acids Research, 2001, 29(9): e45 |
| [10] | Gough NR. Focus issue: recruiting players for a game of ERK[J]. Science Signaling, 2011, 4(196): eg9 |
| [11] | Li HE, Yao WJ, Fu YR, et al. De Novo assembly and discovery of genes that are involved in drought tolerance in Tibetan Sophora moorcroftiana[J]. PLoS One, 2015, 10(1): e111054 |
| [12] | Chen CB, Harel A, Gorovoits R, et al. MAPK regulation of sclerotial development in Sclerotinia sclerotiorum is linked with pH and cAMP sensing[J]. Molecular Plant-Microbe Interactions, 2004, 17(4): 404-413 |